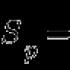Описание на плазмената мембрана. Каква е структурата на плазмената мембрана? Какви са неговите функции
Структурата на клетките на живите организми до голяма степен зависи от това какви функции изпълняват. Въпреки това, има редица архитектурни принципи, общи за всички клетки. По-специално, всяка клетка има обвивка от външната страна, която се нарича цитоплазмена или плазмена мембрана. Има и друго име - плазмолема.
структура
Плазмената мембрана се състои от три основни типа молекули – протеини, въглехидрати и липиди. В различни видовеклетки, съотношението на тези компоненти може да варира.
През 1972 г. учените Никълсън и Сингър предлагат флуидно-мозаичен модел на структурата на цитоплазмената мембрана. Този модел послужи като отговор на въпроса за структурата на клетъчната мембрана и не е загубил своята актуалност и до днес. Същността на флуидно-мозаечния модел е както следва:
- Липидите са подредени в два слоя, формиращи основата на клетъчната стена;
- Хидрофилните краища на липидните молекули са навътре, докато хидрофобните краища са навън;
- Отвътре тази структура има слой от протеини, които проникват в липидите като мозайка;
- В допълнение към протеините тук има малко количество въглехидрати - хексоза;
Това биологична системаима голяма мобилност. Протеиновите молекули могат да се подреждат, фокусирайки се от едната страна на липидния слой, или могат да се движат свободно и да променят позицията си.
Функции
Въпреки някои различия в структурата, плазмолемите на всички клетки имат набор от общи функции. В допълнение, те могат да имат характеристики, които са силно специфични за даден тип клетка. Нека разгледаме накратко общите основни функции на всички клетъчни мембрани:

Селективна пропускливост
Основното свойство на плазмената мембрана е селективната пропускливост. През него преминават йони, аминокиселини, глицерол и мастни киселини, глюкоза. В същото време клетъчната мембрана позволява на някои вещества да преминават и задържа други.
Има няколко вида механизми за транспортиране на вещества през клетъчната мембрана:
- Дифузия;
- Осмоза;
- Екзоцитоза;
- Ендоцитоза;
 Дифузията и осмозата не изискват енергийни разходи и се извършват пасивно, останалите видове транспорт са активни процеси, които консумират енергия.
Дифузията и осмозата не изискват енергийни разходи и се извършват пасивно, останалите видове транспорт са активни процеси, които консумират енергия.
Това свойство на клетъчната мембрана по време на пасивен транспорт се дължи на наличието на специални интегрални протеини. Такива протеинови канали проникват в плазмалемата и образуват пасажи в нея. Калциеви, калиеви и lor йони се движат по такива канали спрямо градиента на концентрация.
Транспорт на вещества
Основните свойства на плазмената мембрана включват и способността й да транспортира молекули от различни вещества.
Описани са следните механизми на пренос на вещества през плазмалемата:
- Пасивни - дифузия и осмоза;
- Активен;
- Транспортиране в мембранна опаковка;
Нека разгледаме тези механизми по-подробно.
Пасивен
![]() Пасивните видове транспорт включват осмоза и дифузия. Дифузията е движение на частици по градиент на концентрация. В този случай клетъчната стена действа като осмотична бариера. Скоростта на дифузия зависи от размера на молекулите и тяхната разтворимост в липиди. Дифузията от своя страна може да бъде неутрална (с пренасяне на незаредени частици) или улеснена, когато са включени специални транспортни протеини.
Пасивните видове транспорт включват осмоза и дифузия. Дифузията е движение на частици по градиент на концентрация. В този случай клетъчната стена действа като осмотична бариера. Скоростта на дифузия зависи от размера на молекулите и тяхната разтворимост в липиди. Дифузията от своя страна може да бъде неутрална (с пренасяне на незаредени частици) или улеснена, когато са включени специални транспортни протеини.
Осмозата е дифузия на водни молекули през клетъчната стена..
Полярните молекули с голяма маса се транспортират с помощта на специални протеини - този процес се нарича улеснена дифузия. Транспортните протеини проникват през клетъчната мембрана и образуват канали. Всички транспортни протеини се разделят на каналообразуващи и транспортери. Проникването на заредени частици се улеснява от наличието на мембранен потенциал.
Активен
![]() Транспортирането на вещества през клетъчната стена срещу електрохимичен градиент се нарича активен транспорт. Такъв транспорт винаги се осъществява с участието на специални протеини и изисква енергия. Транспортните протеини имат специални места, които се свързват с транспортираното вещество. Колкото повече такива сайтове, толкова по-бърз и по-интензивен е трансферът. По време на трансфер на протеинтранспортерът претърпява обратими структурни промени, което му позволява да изпълнява функциите си.
Транспортирането на вещества през клетъчната стена срещу електрохимичен градиент се нарича активен транспорт. Такъв транспорт винаги се осъществява с участието на специални протеини и изисква енергия. Транспортните протеини имат специални места, които се свързват с транспортираното вещество. Колкото повече такива сайтове, толкова по-бърз и по-интензивен е трансферът. По време на трансфер на протеинтранспортерът претърпява обратими структурни промени, което му позволява да изпълнява функциите си.
В мембранна опаковка
Молекули от органични вещества с голяма маса се пренасят през мембраната с образуване на затворени мехурчета - везикули, които образуват мембраната.
отличителен белегвезикуларен транспорт е, че пренесените макрочастици не се смесват с други клетъчни молекули или нейните органели.
Прехвърлянето на големи молекули в клетката се нарича ендоцитоза. От своя страна ендоцитозата се разделя на два вида - пиноцитоза и фагоцитоза. В този случай част от плазмената мембрана на клетката образува мехурче около пренесените частици, наречено вакуола. Размерите на вакуолите по време на пиноцитоза и фагоцитоза имат значителни разлики.
В процеса на пиноцитоза се случва усвояването на течности от клетката. Фагоцитозата осигурява усвояването на големи частици, фрагменти от клетъчни органели и дори микроорганизми.
Екзоцитоза
 Екзоцитозата е отстраняването на вещества от клетката. В този случай вакуолите се придвижват към плазмалемата. Освен това стената на вакуолата и плазмалематазапочват да се слепват и след това се сливат. Веществата, които се съдържат във вакуолата, се придвижват до заобикаляща среда.
Екзоцитозата е отстраняването на вещества от клетката. В този случай вакуолите се придвижват към плазмалемата. Освен това стената на вакуолата и плазмалематазапочват да се слепват и след това се сливат. Веществата, които се съдържат във вакуолата, се придвижват до заобикаляща среда.
Клетки на някои прости организмиимат строго определени зони, за да осигурят такъв процес.
И ендоцитозата, и екзоцитозата протичат в клетката с участието на фибриларните компоненти на цитоплазмата, които имат тясна пряка връзка с плазмената мембрана.
Ядрото е отговорно за съхраняването на генетичния материал, записан върху ДНК, а също така контролира всички процеси на клетката. Цитоплазмата съдържа органели, всяка от които има свои собствени функции, като например синтез на органични вещества, храносмилане и т. н. А за последния компонент ще говорим по-подробно в тази статия.
по биология?
говорене прост език, това е черупка. Въпреки това, не винаги е напълно непроницаем. Транспортирането на определени вещества през мембраната е почти винаги разрешено.
В цитологията мембраните могат да бъдат разделени на два основни типа. Първият е плазмената мембранакойто покрива клетката. Вторият е мембраните на органелите. Има органели, които имат една или две мембрани. Едномембранните клетки включват ендоплазмения ретикулум, вакуоли и лизозоми. Пластидите и митохондриите принадлежат към двумембранните.
Също така, мембраните могат да бъдат вътре в органели. Обикновено това са производни на вътрешната мембрана на двумембранни органели.
Как са подредени мембраните на двумембранните органели?
Пластидите и митохондриите имат две мембрани. Външната мембрана на двете органели е гладка, но вътрешната образува структурите, необходими за функционирането на органоида.
И така, черупката на митохондриите има издатини навътре - кристи или хребети. Те преминават през цикъла химична реакциянеобходими за клетъчното дишане.
Производни на вътрешната мембрана на хлоропластите са дисковидни торбички - тилакоиди. Събрани са на купчини – зърна. Отделните грани се комбинират помежду си с помощта на ламели - дълги структури, също образувани от мембрани.
Структурата на мембраните на едномембранните органели
Тези органели имат само една мембрана. Обикновено това е гладка мембрана, съставена от липиди и протеини.
Характеристики на структурата на плазмената мембрана на клетката
Мембраната е изградена от вещества като липиди и протеини. Структурата на плазмената мембрана осигурява нейната дебелина от 7-11 нанометра. По-голямата част от мембраната е изградена от липиди.

Структурата на плазмената мембрана предвижда наличието на два слоя в нея. Първият е двоен слой от фосфолипиди, а вторият е слой от протеини.
Липиди на плазмената мембрана
Липидите, които изграждат плазмената мембрана, са разделени на три групи: стероиди, сфингофосфолипиди и глицерофосфолипиди. Молекулата на последния съдържа остатък от тривалентния алкохол глицерол, в който водородните атоми на две хидроксилни групи са заменени с вериги на мастни киселини, а водородният атом на третата хидроксилна група е заменен с остатък фосфорна киселина, който от своя страна се присъединява към остатъка от една от азотните основи.
Молекулата на глицерофосфолипида може да бъде разделена на две части: глава и опашка. Главата е хидрофилна (тоест разтваря се във вода), а опашките са хидрофобни (отблъскват водата, но се разтварят в органични разтворители). Поради тази структура, молекулата на глицерофосфолипидите може да се нарече амфифилна, тоест едновременно хидрофобна и хидрофилна.
Сфингофосфолипидите са сходни в химическа структураза глицерофосфолипиди. Но те се различават от споменатите по-горе по това, че в състава си, вместо остатък от глицерол, имат остатък от сфингозин алкохол. Техните молекули също имат глави и опашки.
Снимката по-долу ясно показва структурата на плазмената мембрана.

Протеини на плазмената мембрана
Що се отнася до протеините, които изграждат структурата на плазмената мембрана, това са основно гликопротеини.
В зависимост от местоположението им в черупката те могат да бъдат разделени на две групи: периферни и интегрални. Първите са тези, които са на повърхността на мембраната, а вторите са тези, които проникват в цялата дебелина на мембраната и са вътре в липидния слой.
В зависимост от функциите, които изпълняват протеините, те могат да бъдат разделени на четири групи: ензими, структурни, транспортни и рецепторни.

Всички протеини, които са в структурата на плазмената мембрана, не са химически свързани с фосфолипиди. Следователно те могат да се движат свободно в основния слой на мембраната, да се събират в групи и т. н. Ето защо структурата на плазмената мембрана на клетката не може да се нарече статична. Той е динамичен, тъй като се променя през цялото време.
Каква е ролята на клетъчната мембрана?
Структурата на плазмената мембрана й позволява да се справи с пет функции.
Първият и основен е ограничаването на цитоплазмата. Поради това клетката има постоянна форма и размер. Тази функция се осигурява от факта, че плазмената мембрана е здрава и еластична.
Втората роля е осигуряване. Поради своята еластичност, плазмените мембрани могат да образуват израстъци и гънки в местата си на свързване.
Следващата функция на клетъчната мембрана е транспортната. Осигурява се от специални протеини. Благодарение на тях в клетката могат да се транспортират необходимите вещества, а от нея да се изхвърлят ненужните.

Освен това плазмената мембрана изпълнява ензимна функция. Осъществява се и благодарение на протеините.
И последната функция е сигнализирането. Поради факта, че протеините под влияние на определени условия могат да променят своята пространствена структура, плазмената мембрана може да изпраща сигнали до клетките.
Сега знаете всичко за мембраните: какво е мембрана в биологията, какво представляват, как са подредени плазмената мембрана и органоидните мембрани, какви функции изпълняват.
Лекция
плазмената мембрана
Планирайте
1. Структурата на плазмената мембрана
2. Функции на плазмената мембрана. Механизми на транспортиране на вещества през плазмалемата. Рецепторна функция на плазмалемата
Междуклетъчни контакти
1.
Структурата на плазмената мембрана
плазмената мембрана или плазмалемата,е повърхностна периферна структура, която ограничава клетката отвън и осигурява връзката й с други клетки и извънклетъчната среда. Има дебелина около 10 nm. Сред другите клетъчни мембрани плазмалемата е най-дебелата. Химически плазмената мембрана е липопротеинов комплекс.Основните компоненти са липидите (около 40%), протеините (повече от 60%) и въглехидратите (около 2-10%). Липидите са голяма групаорганични вещества, които имат лоша разтворимост във вода (хидрофобност) и добра разтворимост в органични разтворители и мазнини (липофилност). Представителни липиди, открити в плазмената мембрана, са фосфолипиди, сфингомиелини и холестерол. В растителните клетки холестеролът се заменя с фитостерол. Според тяхната биологична роля плазмалемните протеини могат да се разделят на ензимни протеини, рецепторни и структурни протеини.Въглехидратите на плазмалемата са част от плазмалемата в свързано състояние (гликолипиди и гликопротеини). В момента е общоприето флуидно-мозаичен модел на структурата на биологичната мембрана.Според този модел, структурната основа на мембраната е образувана от двоен слой фосфолипиди, инкрустирани с протеини. Опашките на молекулите са обърнати една към друга в двоен слой, докато полярните глави остават отвън, образувайки хидрофилни повърхности. Протеиновите молекули не образуват непрекъснат слой, те са разположени в липидния слой, потъвайки на различни дълбочини (има периферни протеини, някои протеини проникват през мембраната, други са потопени в липидния слой). Повечето протеини не са свързани с мембранни липиди; те сякаш плуват в "липидно езеро". Следователно, протеиновите молекули могат да се движат по мембраната, да се събират в групи или, обратно, да се разпръскват по повърхността на мембраната. Това предполага, че плазмената мембрана не е статична, замръзнала формация. Извън плазмалемата е епимембранният слой - гликокаликс.
Дебелината на този слой е около 3-4 nm. Гликокаликсът се намира в почти всички животински клетки. Той е свързан с плазмената мембрана гликопротеинов комплекс.Въглехидратите образуват дълги, разклонени вериги от полизахариди, свързани с протеини и липиди на плазмената мембрана. Гликокаликсът може да съдържа ензимни протеини, участващи в извънклетъчното разграждане на различни вещества. Продуктите на ензимната активност (аминокиселини, нуклеотиди, мастни киселини и др.) се транспортират през плазмената мембрана и се абсорбират от клетките. Плазменната мембрана непрекъснато се обновява. Това се случва чрез отделяне на малки мехурчета от нейната повърхност в клетката и вграждане на вакуоли от вътрешността на клетката в мембраната. По този начин в клетката има постоянен поток от мембранни елементи: от плазмената мембрана в цитоплазмата (ендоцитоза)и потока на мембранни структури от цитоплазмата към клетъчната повърхност (екзоцитоза).В циркулацията на мембраните водеща роля се отрежда на системата от мембранни вакуоли на комплекса Голджи. 2. Функции на плазмената мембрана. Механизми на транспортиране на вещества през плазмалемата. Рецепторна функция на плазмалемата Плазменната мембрана изпълнява редица важни функции: 1)
Бариера.Бариерната функция на плазмената мембрана е да ограничи свободната дифузия на вещества от клетка в клетка, да предотврати изтичането на водоразтворимо съдържание на клетката. Но тъй като клетката трябва да получава необходимите хранителни вещества, да освобождава крайните продукти на метаболизма и да регулира вътреклетъчните концентрации на йони, в нея са се формирали специални механизми за пренос на веществата през клетъчната мембрана. 2)
Транспорт.Транспортната функция е
Осигуряване на влизане и излизане на различни вещества в и извън клетката. Важно свойство на мембраната е селективна пропускливост, или полупропускливост.Той лесно пропуска вода и водоразтворими газове и отблъсква полярни молекули като глюкоза или аминокиселини. Има няколко механизма за транспортиране на вещества през мембраната: пасивен транспорт;
активен транспорт;
транспортиране в мембранна опаковка.
Трансферът на полярни молекули (захари, аминокиселини), извършен с помощта на специални мембранни транспортни протеини, се нарича улеснена дифузия.Такива протеини се намират във всички видове биологични мембрани и всеки конкретен протеин е предназначен да носи молекули от определен клас. Транспортните протеини са трансмембранни; тяхната полипептидна верига пресича липидния бислой няколко пъти, образувайки през пасажи в него. Това осигурява пренос на специфични вещества през мембраната без директен контакт с нея. Има два основни класа транспортни протеини: протеини носители (транспортьори)и каналообразуващипротеини (протеинови канали). Протеините носители пренасят молекули през мембраната, като първо променят конфигурацията си. Белтъците, образуващи канали, образуват пълни с вода пори в мембраната. Когато порите са отворени, молекулите на специфични вещества (обикновено неорганични йони правилен размери заряд) преминават през тях. Ако молекулата на транспортираното вещество няма заряд, тогава посоката на транспорт се определя от градиента на концентрация. Ако молекулата е заредена, тогава нейният транспорт, освен градиента на концентрация, се влияе и от електрическия заряд на мембраната (мембранен потенциал). Вътрешна странаПлазмалемата обикновено е отрицателно заредена спрямо външната. Мембранният потенциал улеснява проникването на положително заредени йони в клетката и предотвратява преминаването на отрицателно заредени йони. активен транспорт.Активният транспорт е движението на вещества срещу електрохимичен градиент. То винаги се осъществява от транспортиращи протеини и е тясно свързано с енергиен източник. Протеините носители имат места на свързване с транспортираното вещество. Колкото повече такива сайтове са свързани с веществото, толкова по-висока е скоростта на транспортиране. Селективният трансфер на едно вещество се нарича унипорт.Извършва се пренос на няколко вещества котранспортни системи.Ако трансферът върви в една посока, това е така симпорт,ако е в противоположното антипорт.Например, глюкозата се транспортира от извънклетъчната течност в клетката по еднопортен начин. Трансфер на глюкоза и Na 4от чревната кухина или тубулите на бъбреците, съответно, към чревните клетки или кръвта се извършва симпортално, а преносът на C1~ и HCO" е антипорт. Предполага се, че по време на трансфера настъпват обратими конформационни промени в транспортера, което позволява движението на вещества, свързани с него. Пример за протеин-носител, който използва енергията, освободена по време на хидролизата на АТФ, за транспортиране на вещества е на+
-ДА СЕ+
помпа,намира се в плазмената мембрана на всички клетки. на +-K помпата работи на принципа на антипорт, изпомпвайки Na" от клетката и K т вътре в клетката срещу техните електрохимични градиенти. Na градиент +създава осмотично налягане, поддържа клетъчния обем и осигурява транспорт на захари и аминокиселини. Работата на тази помпа изразходва една трета от цялата енергия, необходима за жизнената дейност на клетките. При изследване на механизма на действие на Na +-К +Установено е, че помпа е АТФазен ензим и трансмембранен интегрален протеин. В присъствието на Na +и АТФ, под действието на АТФаза, крайният фосфат се отделя от АТФ и се прикрепя към остатъка от аспарагинова киселина върху молекулата на АТФаза. Молекулата АТФаза се фосфорилира, променя конфигурацията си и Na +се отстранява от клетката. След отстраняването на Na от клетката винаги се осъществява транспортирането на К" в клетката. За да се направи това, предварително прикрепеният фосфат се отцепва от АТФаза в присъствието на К. Ензимът се дефосфорилира, възстановява конфигурацията си и К 1"напомпани" в клетката. АТФазата се образува от две субединици, голяма и малка. Голямата субединица се състои от хиляди аминокиселинни остатъци, които пресичат бислоя няколко пъти. Той има каталитична активност и може да бъде обратимо фосфорилиран и дефосфорилиран. Голямата субединица от цитоплазмената страна има места за свързване на Na +и АТФ, и навън- K сайтове на свързване +и ouabaina. Малката субединица е гликопротеин и нейната функция все още не е известна. на +-К помпата има електрогенен ефект. Той премахва три положително заредени Na йона е от клетката и вкарва в нея два йона К. В резултат на това през мембраната протича ток, който образува електрически потенциал с отрицателна стойност във вътрешната част на клетката спрямо външната й повърхност. Na"-K +помпата регулира обема на клетките, контролира концентрацията на вещества вътре в клетката, поддържа осмотичното налягане и участва в създаването на мембранен потенциал. Транспортиране в мембранна опаковка.
Пренасянето на макромолекули (протеини, нуклеинови киселини, полизахариди, липопротеини) и други частици през мембраната се осъществява чрез последователно образуване и сливане на везикули (везикули), заобиколени от мембраната. Процесът на везикуларен транспорт протича на два етапа. Първоначално везикулната мембрана и плазмалемата се слепват и след това се сливат. За хода на етап 2 е необходимо водните молекули да бъдат изместени от взаимодействащи липидни бислоеве, които се приближават един до друг на разстояние от 1-5 nm. Смята се, че този процес се активира от специални слети протеини(те са изолирани досега само във вируси). Везикуларен транспорт има важна характеристика- абсорбираните или секретираните макромолекули във везикулите обикновено не се смесват с други макромолекули или органели на клетката. Мехурчетата могат да се сливат със специфични мембрани, което осигурява обмена на макромолекули между извънклетъчното пространство и съдържанието на клетката. По същия начин макромолекулите се прехвърлят от едно клетъчно отделение в друго. Транспортирането на макромолекули и частици в клетката се нарича ендоцитоза.В този случай транспортираните вещества се обвиват от част от плазмената мембрана, образува се мехурче (вакуола), което се движи вътре в клетката. В зависимост от размера на образуваните везикули се разграничават два вида ендоцитоза - пиноцитоза и фагоцитоза.
пиноцитозаосигурява абсорбция на течни и разтворени вещества под формата на малки мехурчета (d=150 nm). Фагоцитоза -това е усвояването на големи частици, микроорганизми или фрагменти от органели, клетки. В този случай се образуват големи везикули, фагозоми или вакуоли (d-250 nm или повече). При протозоите фагоцитната функция е форма на хранене. При бозайниците фагоцитната функция се осъществява от макрофаги и неутрофили, които предпазват тялото от инфекция чрез поглъщане на нахлуващи микроби. Макрофагите също участват в изхвърлянето на стари или увредени клетки и техните фрагменти (в човешкото тяло макрофагите абсорбират повече от 100 стари червени кръвни клетки дневно). Фагоцитозата започва едва когато абсорбираната частица се свърже с повърхността на фагоцита и активира специализирани рецепторни клетки. Свързването на частиците със специфични мембранни рецептори причинява образуването на псевдоподии, които обгръщат частицата и, сливайки се по краищата, образуват мехурче - фагозома.Образуването на фагозома и правилната фагоцитоза се случва само ако по време на процеса на обвиване, частицата е в постоянен контакт с рецепторите на плазмалемата, сякаш се „закопчава“. Значителна част от материала, абсорбиран от клетката чрез ендоцитоза, завършва в лизозоми. Големи частици са включени в фагозомикоито след това се сливат с лизозоми, за да се образуват фаголизозоми.Течността и макромолекулите, поети по време на пиноцитозата, първоначално се прехвърлят към ендозоми, които също се сливат с лизозоми, за да образуват ендолизозоми. Различни хидролитични ензими, присъстващи в лизозомите, бързо разрушават макромолекулите. Продуктите на хидролизата (аминокиселини, захари, нуклеотиди) се транспортират от лизозомите до цитозола, където се използват от клетката. Повечето от мембранните компоненти на ендоцитните везикули от фагозоми и ендозоми се връщат чрез екзоцитоза в плазмената мембрана и се използват повторно там. Основното биологично значение на ендоцитозата е придобиването на градивни елементи чрез вътреклетъчно смилане на макромолекулите в лизозомите. Усвояването на веществата в еукариотните клетки започва в специализирани зони на плазмената мембрана, т.нар. оградени ями.На електронни микрографии ямките изглеждат като инвагинации на плазмената мембрана, чиято цитоплазмена страна е покрита с фиброзен слой. Слоят сякаш граничи с малки ями на плазмалемата. Ямките заемат около 2% от общата повърхност на еукариотната клетъчна мембрана. В рамките на една минута ямките нарастват, инвагинират все по-дълбоко и по-дълбоко, привличат се в клетката и след това, стеснявайки се в основата, се отцепват, образувайки оградени везикули. Установено е, че приблизително една четвърт от мембраната под формата на граничещи везикули се отцепва от плазмената мембрана на фибробластите в рамките на една минута. Везикулите бързо губят границата си и придобиват способността да се сливат с лизозомата. Ендоцитоза може да бъде неспецифични(конститутивно) и специфични(рецептор). В неспецифична ендоцитозаклетката улавя и абсорбира вещества, които са й напълно чужди, например частици сажди, багрила. Първоначално частиците се отлагат върху гликокаликса на плазмалемата. Положително заредените протеинови групи са особено добре утаени (адсорбирани), тъй като гликокаликсът носи отрицателен заряд. Тогава морфологията на клетъчната мембрана се променя. Той може или да потъне, образувайки инвагинации (инвагинации), или, обратно, да образува израстъци, които сякаш се сгъват, отделяйки малки обеми от течната среда. Образуването на инвагинации е по-характерно за клетките на чревния епител, амебите, а израстъците - за фагоцитите и фибробластите. Тези процеси могат да бъдат блокирани от респираторни инхибитори. Получените везикули - първични ендозоми - могат да се слеят помежду си, увеличавайки се по размер. Впоследствие те се комбинират с лизозоми, превръщайки се в ендолизозома - храносмилателна вакуола. Интензивността на течнофазовата неспецифична пиноцитоза е доста висока. Макрофагите образуват до 125, а епителните клетки на тънките черва до хиляда пинозоми в минута. Изобилието от пиносоми води до факта, че плазмалемата бързо се изразходва за образуването на много малки вакуоли. Възстановяването на мембраната протича доста бързо по време на рециклиране по време на екзоцитоза поради връщането на вакуоли и включването им в плазмалемата. При макрофагите цялата плазмена мембрана се заменя за 30 минути, а във фибробластите за 2 часа. | Повече ▼ ефективен начинабсорбцията от извънклетъчната течност на специфични макромолекули е специфична ендоцитоза(медиирано от рецептори). В този случай макромолекулите се свързват с комплементарни рецептори на клетъчната повърхност, натрупват се в оградената ямка и след това, образувайки ендозома, се потапят в цитозола. Рецепторната ендоцитоза осигурява натрупването на специфични макромолекули в неговия рецептор. Молекулите, които се свързват с рецептор на повърхността на плазмалемата, се наричат лиганди.С помощта на рецепторна ендоцитоза в много животински клетки холестеролът се абсорбира от извънклетъчната среда. Плазменната мембрана участва в отстраняването на веществата от клетката (екзоцитоза). В този случай вакуолите се приближават до плазмалемата. В точките на контакт плазмолемата и вакуолната мембрана се сливат и съдържанието на вакуолата навлиза в околната среда. При някои протозои местата на клетъчната мембрана за екзоцитоза са предварително определени. Така че в плазмената мембрана на някои цилиарни реснички има определени области с правилно местоположениеголеми глобули от интегрални протеини. Мукоцистите и трихоцистите на ресничките, които са напълно готови за секреция, имат ореол от интегрални белтъчни глобули в горната част на плазмалемата. Тези участъци от мембраната на мукоцистите и трихоцистите са в контакт с повърхността на клетката. При неутрофилите се наблюдава особена екзоцитоза. Те са в състояние, при определени условия, да освободят своите лизозоми в околната среда. В някои случаи се образуват малки израстъци на плазмалемата, съдържащи лизозоми, които след това се откъсват и преминават в околната среда. В други случаи има инвагинация на плазмалемата дълбоко в клетката и нейното улавяне на лизозоми, разположени далеч от повърхността на клетката. Процесите на ендоцитоза и екзоцитоза се осъществяват с участието на системата от фибриларни компоненти на цитоплазмата, свързани с плазмолемата. Рецепторна функция на плазмалемата.Този от основната, универсална за всички клетки, е рецепторната функция на плазмалемата. Той определя взаимодействието на клетките помежду си и с външна среда..
Цялото разнообразие от информационни междуклетъчни взаимодействия може да бъде схематично представено като верига от последователни реакции сигнал-рецептор-вторичен пратеник-отговор (концепция сигнал-отговор).Прехвърлянето на информация от клетка към клетка се осъществява чрез сигнални молекули, които се произвеждат в някои клетки и специфично засягат други, които са чувствителни към сигнала (целеви клетки). Сигнална молекула - първичен посредниксе свързва с рецептори, разположени върху целевите клетки, които реагират само на определени сигнали. Сигнални молекули - лиганди -се приближават до рецептора си като ключ към ключалка. Лиганди за мембранни рецептори (плазмалема рецептори) са хидрофилни молекули, пептидни хормони, невротрансмитери, цитокини, антитела, а за ядрени рецептори - мастноразтворими молекули, стероидни и тироидни хормони, витамин D. Мембранните протеини или гликокаликсните елементи могат да действат като рецептори върху клетъчна повърхност - полизахариди и гликопротеини. Смята се, че областите, чувствителни към отделни вещества, са разпръснати по повърхността на клетката или са събрани в малки зони. И така, на повърхността на прокариотните клетки и животинските клетки има ограничен брой места, с които вирусните частици могат да се свързват. Мембранните протеини (носители и канали) разпознават, взаимодействат и пренасят само определени вещества. Клетъчните рецептори участват в предаването на сигнали от повърхността на клетката в нея. Разнообразието и спецификата на наборите от рецептори на клетъчната повърхност води до създаването на много сложна системамаркери, които разграничават себе си от не-аз клетките. Подобни клетки взаимодействат помежду си, техните повърхности могат да се слепват (конюгиране в протозои, образуване на тъкан в многоклетъчни). Клетките, които не възприемат маркери, както и тези, които се различават по набора от детерминантни маркери, се унищожават или отхвърлят. Когато се образува комплексът рецептор-лиганд, се активират трансмембранни протеини: конвертор протеин, усилвателен протеин. В резултат на това рецепторът променя своята конформация и взаимодейства с предшественика на втория пратеник, разположен в клетката - пратеник.Пратениците могат да бъдат йонизиран калций, фосфолипаза С, аденилатциклаза, гуанилатциклаза. Под въздействието на месинджъра, активирането на ензими, участващи в синтеза циклични монофосфати - AMPили HMF.Последните променят активността на два вида ензими протеин киназа в клетъчната цитоплазма, което води до фосфорилиране на множество вътреклетъчни протеини. Най-честото образуване на cAMP, под въздействието на което се повишава секрецията на редица хормони - тироксин, кортизон, прогестерон, разграждането на гликогена в черния дроб и мускулите, честотата и силата на сърдечните контракции, остеодеструкцията и обратното абсорбцията на вода в нефронните тубули се увеличава. Активността на аденилатциклазната система е много висока – синтезът на сАМР води до десетхилядно увеличение на сигнала. Под действието на cGMP секрецията на инсулин от панкреаса, хистамин от мастоцитите, серотонин от тромбоцитите се увеличава и гладката мускулна тъкан се намалява. В много случаи образуването на комплекс рецептор-лиганд води до промяна в мембранния потенциал, което от своя страна води до промяна в пропускливостта на плазмалемата и метаболитните процеси в клетката. 3.
Междуклетъчни контакти липопротеинов рецептор на плазмената мембрана При многоклетъчните животни в образуването участва плазмолемата междуклетъчни връзкиосигуряване на междуклетъчни взаимодействия. Има няколко вида такива структури. § Прост контакт.Прост контакт се открива сред повечето клетки от различен произход, съседни една на друга.
Представлява конвергенцията на плазмените мембрани на съседни клетки на разстояние 15-20 nm. В този случай възниква взаимодействието на гликокаликсните слоеве на съседните клетки. § Тесен (затварящ) контакт.При такава връзка външните слоеве на двете плазмени мембрани са максимално близки. Сближаването е толкова плътно, че има един вид сливане на участъци от плазмените мембрани на две съседни клетки. Сливането на мембраните не се случва по цялата площ на плътен контакт, а е поредица от точкова конвергенция на мембраните. Ролята на плътния контакт е механично да свързва клетките една с друга. Тази област е непроницаема за макромолекули и йони и следователно блокира, ограничава междуклетъчните празнини (и заедно с тях всъщност вътрешна средаорганизъм) от външната среда. § Петното на сцепление или десмозома.Дезмозомата е малка площ до 0,5 µm в диаметър. В зоната на десмозомата от страната на цитоплазмата има зона от тънки фибрили. Функционалната роля на десмозомите е главно в механичната връзка между клетките. § Празнина на контакт или нексус.При този тип контакт плазмените мембрани на съседните клетки са разделени с празнина от 2-3 nm на разстояние 0,5-3 µm. В структурата на плазмолемите са разположени специални протеинови комплекси (конексони). Един конексон на плазмената мембрана на клетката е точно противопоставен от конексон на плазмената мембрана на съседната клетка. В резултат на това се образува канал от една клетка в друга. Connexons могат да се свиват, променяйки диаметъра на вътрешния канал и по този начин да участват в регулирането на транспорта на молекули между клетките. Този тип връзка се среща във всички групи тъкани. Функционалната роля на междинната връзка е да пренася йони и малки молекули от клетка в клетка. И така, в сърдечния мускул възбуждането, което се основава на процеса на промяна на йонната пропускливост, се предава от клетка на клетка през нексуса. § Синаптичен контакт или синапс.Синапсите са зони на контакт между две клетки, специализирани за еднопосочно предаване на възбуждане или инхибиране от един елемент на друг. Този тип връзка е характерна за нервната тъкан и се среща както между два неврона, така и между неврон и друг елемент. Мембраните на тези клетки са разделени от междуклетъчно пространство – синаптична цепнатина с ширина около 20-30 nm. Мембраната в зоната на синаптичен контакт на една клетка се нарича пресинаптична, а другата - постсинаптична. В близост до пресинаптичната мембрана страхотно количествомалки вакуоли (синаптични везикули), съдържащи невротрансмитер. В момента на преминаване на нервния импулс синаптичните везикули изхвърлят невротрансмитера в синаптичната цепнатина. Медиаторът взаимодейства с рецепторните места на постсинаптичната мембрана, което в крайна сметка води до предаване на нервен импулс. В допълнение към предаването на нервен импулс, синапсите осигуряват твърда връзка между повърхностите на две взаимодействащи клетки. § Плазмодесма.Този тип междуклетъчна комуникация се среща в растенията. Плазмодесмите са тънки тръбести канали, които свързват две съседни клетки. Диаметърът на тези канали обикновено е 40-50 nm. Плазмодесмите преминават през клетъчната стена, която разделя клетките. В младите клетки броят на плазмодесмите може да бъде много висок (до 1000 на клетка). С остаряването на клетките броят им намалява поради разкъсвания с увеличаване на дебелината на клетъчната стена. Функционалната роля на плазмодесмата е да осигури междуклетъчната циркулация на разтвори, съдържащи хранителни вещества, йони и други съединения. Plasmodesmata инфектират клетките с растителни вируси. Специализирани структури на плазмената мембрана Плазмалемата на много животински клетки образува израстъци от различни структури (микровили, реснички, флагели). Най-често се намира на повърхността на много животински клетки микровили.Тези израстъци на цитоплазмата, ограничени от плазмалемата, имат формата на цилиндър със заоблен връх. Микровилите са характерни за епителните клетки, но се срещат и в клетките на други тъкани. Микровилите са с диаметър около 100 nm. Техният брой и дължина са различни при различните типове клетки. Значението на микровилите се състои в значително увеличаване на площта на клетъчната повърхност. Това е особено важно за клетките, участващи в усвояването. Така че, в чревния епител на 1 мм 2повърхности, има до 2x10 8 микровили.
Мембраната се състои от два слоя липидни молекули (двуслойни), в които са вградени протеини. Въглехидратите са прикрепени към някои липидни и протеинови молекули. Малко са от тях. Дебелината на мембраната е около 10 nm (0,00001 mm). Основната част от мембраната е непрекъснат слой от фосфолипидни молекули. В това течностпотопени в слой молекули с различна структура и функции на протеинови молекули. Протеините не покриват напълно липидния бислой, а са разположени в него поотделно или на групи. Като цяло това напомня мозайка(фиг. 2. B. C). В тази връзка се нарича приетият в момента мембранен модел течна мозайка. Протеините са способни ходпо протежение на липидния слой. Молекулите на липидния слой също се движат. Ясно е, че движението на мембранните молекули променя физикохимичните характеристики на последните, а това от своя страна променя функционални характеристикимембрани. Трябва да се отбележи, че плазмената мембрана на повечето клетки няма формата на идеална топка. Напротив, има много издатини, вдлъбнатини, които постоянно променят формата и размера си. Получено в напоследъкРезултатите са направили някои корекции в теорията за структурата на мембраните. Показано е, че не всички мембранни протеини са способни на движение и някои части от мембраните се различават по своята структура от класическия липиден бислой.
Фосфолипидната молекула изглежда като глава с две опашки (фиг. 2 А). Главата (глицеролът) е разтворим във вода, хидрофилен, опашките (мастните киселини) са неразтворими във вода, хидрофобни. Следователно, като са във вода, молекули спонтаннозаемат определено положение спрямо водната фаза. Започвайки от водните молекули, опашките са разположени дълбоко в липидния слой, а водоразтворимите глави са обърнати към външната и вътрешната водна среда (фиг. 2. Б). Липидният двуслой се нарича матрикс. Особено внимание заслужава наличието на липиди в клетъчните мембрани, чиято опашка съдържа мастни киселини, които имат двойни връзки в структурата си, разположени през CH 2 групата (-CH = CH - CH = CH - CH -). Такива мастни киселини се наричат ненаситени. Тези киселини са най-силно засегнати от реактивните кислородни видове (ROS), които постоянно присъстват в тялото на всички живи същества. Техният брой особено се увеличава при различни заболявания. Това може да доведе до неблагоприятни последици, които ще обсъдим по-долу.
Протеините, разпръснати в матрицата (фиг. 2в), са подредени по различни начини. Някои от вътрешната и външната повърхност на липидния слой са мембранни или повърхностни протеини, други са полупотопени в мембраната - полуинтегрални протеини, а трети проникват в цялата мембрана - интегрални протеини. Обикновено полуинтегралните и интегралните протеини се комбинират под един термин - вътрешни протеини, тъй като е трудно да се разграничат един от друг. Интегралните протеини най-често се намират в мембраните. Те могат да бъдат представени като единична молекула и да изпълняват всяка една функция или като група (ансамбъл) от протеини. Всеки член на ансамбъла изпълнява строго определена роля. Тези комплекси изпълняват и една или повече крайни функции.
Ориз. 2. Схематична структура на плазмената мембрана.
Трябва да се отбележи, че функциониращите в мембраната интегрални и повърхностни протеини-ензими доста често променят позицията си. В някои случаи е трудно да се определи към какъв тип (повърхностен или интегрален) принадлежи даден мембранен протеин. Например ензимът фосфолипаза А, като се отлага върху мембраната, е повърхностен протеин, но след това се активира, превръща се в интегрален протеин и взаимодействайки с двуслойните липиди, образува от тях арахидонова киселина (фиг. 3). Последният напуска мембраната и се превръща в други активни съединения, които участват в развитието на различни патологични процеси.
Na + , K + - ATP-ase Adrenoreceptor Аденилат циклаза
 Ca 2+ -АТФаза
Ca 2+ -АТФаза
Фосфолипаза
Na + , K + Ca 2+ Арахидонова
кисел G-протеин гликоген
Ориз. 3. Хипотетична схема на локализация на някои мембранни протеини
Напротив, протеините, участващи в движението на веществата през мембраната - например протеини, участващи в улеснената дифузия на Na + , K + -АТФаза или Са 2+ -АТФаза, като правило, не променят позицията си, функционирайки като интегрални протеини (фиг. 3). И накрая, както вече казахме, мембраната може да съдържа сложни комплекси от няколко протеина, свързани в един ансамбъл от една задача. Такива комплекси включват протеини, участващи в предаването на информационен сигнал през мембраната (фиг. 3). Последните включват комплекс, съдържащ три протеина - адренорецептор, G-протеин и аденилатциклаза. Всички тези протеини са от съществено значение за нормалното функциониране на клетката и при патологията. Ще говорим за това по-долу.
Освен липиди и протеини в мембраната има въглехидрати, но те не са разположени като самостоятелни компоненти, а като компоненти на липиди (гликолипиди) или протеини (гликопротеини). Въглехидратите са разположени на външната повърхност на плазмалемата.
Клетъчната мембрана (плазмената мембрана) е тънка, полупропусклива мембрана, която обгражда клетките.
Функция и роля на клетъчната мембрана
Неговата функция е да защитава целостта на вътрешността, като пропуска някои основни вещества в клетката и предотвратява навлизането на други.
Той също така служи като основа за привързаност към някои организми и към други. Така плазмената мембрана осигурява и формата на клетката. Друга функция на мембраната е да регулира растежа на клетките чрез баланс и.
При ендоцитозата липидите и протеините се отстраняват от клетъчната мембрана, докато веществата се абсорбират. При екзоцитоза везикулите, съдържащи липиди и протеини, се сливат с клетъчната мембрана, увеличавайки размера на клетката. , а гъбичните клетки имат плазмени мембрани. Вътрешните, например, също са затворени в защитни мембрани.
Структура на клетъчната мембрана
Плазменната мембрана се състои главно от смес от протеини и липиди. В зависимост от местоположението и ролята на мембраната в тялото, липидите могат да съставляват от 20 до 80 процента от мембраната, като останалата част са протеини. Докато липидите помагат да се направи мембраната гъвкава, протеините контролират и поддържат химичен съставклетки и също така помагат при транспортирането на молекули през мембраната.
Мембранни липиди
Фосфолипидите са основният компонент на плазмените мембрани. Те образуват липиден бислой, в който хидрофилните (привлечени от вода) "глави" региони спонтанно се организират, за да устоят на водния цитозол и извънклетъчната течност, докато хидрофобните (водоотблъскващи) "опашки" участъци са обърнати настрани от цитозола и извънклетъчната течност. Липидният двуслой е полупропусклив, позволявайки само на някои молекули да дифундират през мембраната.
Холестеролът е друг липиден компонент на мембраните на животинските клетки. Молекулите на холестерола са селективно диспергирани между мембранните фосфолипиди. Това помага да се поддържат твърдостта на клетъчните мембрани, като предотвратява твърде плътното опаковане на фосфолипидите. Холестеролът отсъства в мембраните на растителните клетки.
Гликолипидите са разположени на външната повърхност на клетъчните мембрани и са свързани с тях чрез въглехидратна верига. Те помагат на клетката да разпознава други клетки в тялото.
Мембранни протеини
Клетъчната мембрана съдържа два вида свързани протеини. Белтъците на периферната мембрана са външни и са свързани с нея чрез взаимодействие с други протеини. Интегралните мембранни протеини се въвеждат в мембраната и повечето преминават през нея. Части от тези трансмембранни протеини са разположени от двете му страни.
Протеините на плазмената мембрана имат редица различни функции. Структурните протеини осигуряват подкрепа и форма на клетките. Мембранните рецепторни протеини помагат на клетките да комуникират с външната си среда чрез използването на хормони, невротрансмитери и други сигнални молекули. Транспортните протеини, като глобуларни протеини, пренасят молекули през клетъчните мембрани чрез улеснена дифузия. Гликопротеините имат въглехидратна верига, прикрепена към тях. Те са вградени в клетъчната мембрана, подпомагайки обмена и транспорта на молекули.
Органелни мембрани
Някои клетъчни органели също са заобиколени от защитни мембрани. ядро,