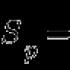слой от клетъчната мембрана. Клетъчни мембрани: тяхната структура и функции. Функции на клетъчната или плазмената мембрана
Цитоплазма- задължителна част от клетката, затворена между плазмената мембрана и ядрото; Подразделя се на хиалоплазма (основното вещество на цитоплазмата), органели (постоянни компоненти на цитоплазмата) и включвания (временни компоненти на цитоплазмата). Химичен съставцитоплазма: основата е вода (60-90% от общата маса на цитоплазмата), различни органични и неорганични съединения. Цитоплазмата е алкална. Отличителен белегцитоплазма на еукариотни клетки - постоянно движение ( циклоза). Открива се основно чрез движението на клетъчни органели, като хлоропласти. Ако движението на цитоплазмата спре, клетката умира, тъй като само в постоянно движение тя може да изпълнява функциите си.
хиалоплазма ( цитозол) е безцветен, лигав, гъст и прозрачен колоиден разтвор. Именно в него протичат всички метаболитни процеси, осигурява взаимовръзката на ядрото и всички органели. В зависимост от преобладаването на течната част или големите молекули в хиалоплазмата се разграничават две форми на хиалоплазма: сол- по-течна хиалоплазма и гел- по-плътна хиалоплазма. Възможни са взаимни преходи между тях: гелът се превръща в зол и обратно.
Функции на цитоплазмата:
- интегриране на всички компоненти на клетката в една система,
- среда за протичане на много биохимични и физиологични процеси,
- среда за съществуване и функциониране на органели.
Клетъчни стени
Клетъчни стениограничаване на еукариотните клетки. Във всяка клетъчна мембрана могат да се разграничат най-малко два слоя. Вътрешният слой е в непосредствена близост до цитоплазмата и е представен от плазмената мембрана(синоними - плазмалема, клетъчна мембрана, цитоплазмена мембрана), върху която се образува външният слой. В животинска клетка тя е тънка и се нарича гликокаликс(образувани от гликопротеини, гликолипиди, липопротеини), в растителна клетка - плътни, т.нар. клетъчна стена(образувани от целулоза).
Всички биологични мембрани имат общи структурни характеристики и свойства. В момента общоприет флуиден мозаечен модел на структурата на мембраната. Основата на мембраната е липиден бислой, образуван главно от фосфолипиди. Фосфолипидите са триглицериди, в които един остатък от мастна киселина е заменен с остатък фосфорна киселина; участъкът от молекулата, в който се намира остатъкът от фосфорна киселина, се нарича хидрофилна глава, участъците, в които се намират остатъци от мастни киселини, се наричат хидрофобни опашки. В мембраната фосфолипидите са подредени по строго подреден начин: хидрофобните опашки на молекулите са обърнати една към друга, а хидрофилните глави са обърнати навън, към водата.
В допълнение към липидите, мембраната съдържа протеини (средно ≈ 60%). Те определят повечето от специфичните функции на мембраната (транспорт на определени молекули, катализа на реакциите, получаване и преобразуване на сигнали от заобикаляща средаи т.н.). Разграничаване: 1) периферни протеини(разположен на външната или вътрешната повърхност на липидния двуслой), 2) полуинтегрални протеини(потопен в липидния двуслой на различна дълбочина), 3) интегрални или трансмембранни протеини(проникват през мембраната през и през, докато са в контакт както с външната, така и с вътрешна средаклетки). Интегралните протеини в някои случаи се наричат каналообразуващи или канални, тъй като те могат да се разглеждат като хидрофилни канали, през които полярните молекули преминават в клетката (липидният компонент на мембраната не би ги пропуснал).

А - хидрофилна глава на фосфолипида; C, хидрофобни опашки на фосфолипида; 1 - хидрофобни области на протеини Е и F; 2, хидрофилни области на протеин F; 3 - разклонена олигозахаридна верига, прикрепена към липид в гликолипидна молекула (гликолипидите са по-рядко срещани от гликопротеините); 4 - разклонена олигозахаридна верига, прикрепена към протеин в гликопротеинова молекула; 5 - хидрофилен канал (функционира като пора, през която могат да преминават йони и някои полярни молекули).
Мембраната може да съдържа въглехидрати (до 10%). Въглехидратният компонент на мембраните е представен от олигозахаридни или полизахаридни вериги, свързани с протеинови молекули (гликопротеини) или липиди (гликолипиди). По принцип въглехидратите са разположени на външната повърхност на мембраната. Въглехидратите осигуряват рецепторните функции на мембраната. В животинските клетки гликопротеините образуват епимембранен комплекс, гликокаликса, с дебелина няколко десетки нанометра. В него са разположени много клетъчни рецептори, с негова помощ се осъществява клетъчна адхезия.
Молекулите на протеини, въглехидрати и липиди са подвижни, способни да се движат в равнината на мембраната. Дебелина плазмената мембрана- приблизително 7,5 nm.
Функции на мембраната
Мембраните изпълняват следните функции:
- отделяне на клетъчно съдържание от външна среда,
- регулиране на метаболизма между клетката и околната среда,
- разделяне на клетката на отделения ("отделения"),
- местоположение на "ензимни конвейери",
- осигуряване на комуникация между клетките в тъканите на многоклетъчни организми (адхезия),
- разпознаване на сигнал.
Най-важните мембранно свойство- селективна пропускливост, т.е. мембраните са силно пропускливи за някои вещества или молекули и слабо пропускливи (или напълно непропускливи) за други. Това свойство е в основата на регулаторната функция на мембраните, която осигурява обмена на вещества между клетката и външната среда. Процесът, при който веществата преминават през клетъчната мембрана, се нарича транспорт на вещества. Разграничаване: 1) пасивен транспорт- процесът на преминаване на вещества, преминаващи без енергия; 2) активен транспорт- процесът на преминаване на вещества, протичащ с цената на енергията.
В пасивен транспортвеществата се придвижват от зона с по-висока концентрация към зона с по-ниска, т.е. по градиента на концентрация. Във всеки разтвор има молекули на разтворителя и разтвореното вещество. Процесът на движение на молекулите на разтвореното вещество се нарича дифузия, движението на молекулите на разтворителя се нарича осмоза. Ако молекулата е заредена, тогава нейният транспорт се влияе от електрическия градиент. Следователно често се говори за електрохимичен градиент, комбиниращ двата градиента заедно. Скоростта на транспортиране зависи от големината на наклона.
Могат да се разграничат следните видове пасивен транспорт: 1) проста дифузия- транспортиране на вещества директно през липидния бислой (кислород, въглероден диоксид); 2) дифузия през мембранни канали- транспорт чрез каналообразуващи протеини (Na +, K +, Ca 2+, Cl -); 3) улеснена дифузия- транспорт на вещества с помощта на специални транспортни протеини, всеки от които е отговорен за движението на определени молекули или групи от свързани молекули (глюкоза, аминокиселини, нуклеотиди); 4) осмоза— транспорт на водни молекули (всички биологични системиразтворителят е вода).
Трябва активен транспортвъзниква, когато е необходимо да се осигури пренос на молекули през мембраната срещу електрохимичния градиент. Този транспорт се осъществява от специални белтъци носители, чиято дейност изисква енергийни разходи. Източникът на енергия са АТФ молекули. Активният транспорт включва: 1) Na + /K + -помпа (натриево-калиева помпа), 2) ендоцитоза, 3) екзоцитоза.

Работете Na + /K + -помпа. За нормално функциониране клетката трябва да поддържа определено съотношение на йони K + и Na + в цитоплазмата и във външната среда. Концентрацията на K + вътре в клетката трябва да бъде значително по-висока, отколкото извън нея, и Na + - обратно. Трябва да се отбележи, че Na + и K + могат свободно да дифундират през порите на мембраната. Na+/K+ помпата противодейства на изравняването на тези йони и активно изпомпва Na+ от клетката и K+ в клетката. Na + /K + -помпата е трансмембранен протеин, способен на конформационни промени, така че да може да прикрепи както K+, така и Na+. Цикълът на работа на Na + /K + -помпата може да бъде разделен на следните фази: 1) прикрепване на Na + от вътрешната страна на мембраната, 2) фосфорилиране на протеина на помпата, 3) освобождаване на Na + в извънклетъчно пространство, 4) прикрепване на К + с навънмембрани, 5) дефосфорилиране на помпения протеин, 6) освобождаване на К + във вътреклетъчното пространство. Натриево-калиевата помпа изразходва почти една трета от цялата енергия, необходима за живота на клетката. По време на един цикъл на работа помпата изпомпва 3Na + от клетката и изпомпва в 2K +.
Ендоцитоза- процесът на усвояване от клетката на големи частици и макромолекули. Има два вида ендоцитоза: 1) фагоцитоза- улавяне и усвояване на големи частици (клетки, клетъчни части, макромолекули) и 2) пиноцитоза- улавяне и усвояване течен материал(разтвор, колоиден разтвор, суспензия). Феноменът фагоцитоза е открит от I.I. Мечников през 1882 г. По време на ендоцитозата плазмената мембрана образува инвагинация, краищата й се сливат и структури, отделени от цитоплазмата с една мембрана, се връзват в цитоплазмата. Много протозои и някои левкоцити са способни на фагоцитоза. Пиноцитоза се наблюдава в епителните клетки на червата, в ендотела на кръвоносните капиляри.
Екзоцитоза- обратният процес на ендоцитозата: отстраняването на различни вещества от клетката. По време на екзоцитозата везикулната мембрана се слива с външната цитоплазмена мембрана, съдържанието на везикулата се отстранява извън клетката и нейната мембрана се включва във външната цитоплазмена мембрана. По този начин хормоните се отделят от клетките на жлезите с вътрешна секреция, а при протозоите остава несмляна храна.
Отидете на лекции номер 5„Клетъчната теория. Видове клетъчна организация»
Отидете на лекции номер 7"Еукариотна клетка: структура и функции на органелите"
Таблица номер 2
Въпрос 1 (8)
клетъчната мембрана(или цитолема, или плазмалема, или плазмена мембрана) отделя съдържанието на всяка клетка от външната среда, като гарантира нейната цялост; регулира обмена между клетката и околната среда; вътреклетъчните мембрани разделят клетката на специализирани затворени отделения - компартменти или органели, в които се поддържат определени условия на околната среда.
Функции на клетъчната или плазмената мембрана
Мембраната осигурява:
1) Селективно проникване в и извън клетката на молекули и йони, необходими за изпълнение на специфични клетъчни функции;
2) Селективен транспорт на йони през мембраната, поддържащ трансмембранна разлика в електрическия потенциал;
3) Спецификата на междуклетъчните контакти.
Поради наличието в мембраната на множество рецептори, които възприемат химически сигнали - хормони, медиатори и др. активни вещества, той е в състояние да промени метаболитната активност на клетката. Мембраните осигуряват специфичността на имунните прояви поради наличието на антигени върху тях - структури, които предизвикват образуването на антитела, които могат специфично да се свържат с тези антигени.
Ядрото и органелите на клетката също са отделени от цитоплазмата чрез мембрани, които предотвратяват свободното движение на водата и разтворените в нея вещества от цитоплазмата към тях и обратно. Това създава условия за разделяне на биохимичните процеси, протичащи в различни отделения (компартименти) вътре в клетката.
структура на клетъчната мембрана
клетъчната мембрана- еластична структура, дебелина от 7 до 11 nm (фиг. 1.1). Състои се основно от липиди и протеини. От 40 до 90% от всички липиди са фосфолипиди - фосфатидилхолин, фосфатидилетаноламин, фосфатидилсерин, сфингомиелин и фосфатидилинозитол. Важен компонент на мембраната са гликолипидите, представени от церебризиди, сулфатиди, ганглиозиди и холестерол.
Основната структура на клетъчната мембранапредставлява двоен слой от фосфолипидни молекули. Поради хидрофобни взаимодействия, въглехидратните вериги на липидните молекули се държат близо една до друга в разширено състояние. Групи от фосфолипидни молекули от двата слоя взаимодействат с протеинови молекули, потопени в липидната мембрана. Поради факта, че повечето от липидните компоненти на бислоя са в течно състояние, мембраната има подвижност и вълнообразна. Неговите участъци, както и протеините, потопени в липидния двуслой, ще се смесват от една част в друга. Подвижността (течността) на клетъчните мембрани улеснява транспортирането на вещества през мембраната.
протеини на клетъчната мембранапредставени главно от гликопротеини.
Разграничаване
интегрални протеинипроникващи през цялата дебелина на мембраната и
периферни протеиниприкрепен само към повърхността на мембраната, главно към нейната вътрешна част.
Периферни протеинипочти всички функционират като ензими (ацетилхолинестераза, киселинни и алкални фосфатази и др.). Но някои ензими са представени и от интегрални протеини - АТФаза.
интегрални протеиниосигуряват селективен обмен на йони през мембранните канали между извънклетъчната и вътреклетъчната течност, а също така действат като протеини - носители на големи молекули.
Мембранните рецептори и антигени могат да бъдат представени както от интегрални, така и от периферни протеини.
Протеините, съседни на мембраната от цитоплазмената страна, принадлежат към клетъчен цитоскелет. Те могат да се прикрепят към мембранните протеини.
Така, протеинова лента 3(номер на лентата по време на белтъчна електрофореза) на еритроцитните мембрани се комбинира в ансамбъл с други молекули на цитоскелета - спектрин чрез нискомолекулен протеин анкирин
Спектрине основният протеин на цитоскелета, съставляващ двуизмерна мрежа, към която е прикрепен актинът.
актинобразува микрофиламенти, които са контрактилния апарат на цитоскелета.
цитоскелетпозволява на клетката да проявява гъвкаво еластични свойства, осигурява допълнителна здравина на мембраната.
Повечето интегрални протеини са гликопротеини. Тяхната въглехидратна част стърчи от клетъчната мембрана навън. Много гликопротеини имат голям отрицателен заряд поради значителното съдържание на сиалова киселина (например молекулата гликофорин). Това осигурява на повърхността на повечето клетки отрицателен заряд, като помага за отблъскване на други отрицателно заредени обекти. Въглехидратните издатини на гликопротеините носят антигени на кръвната група, други антигенни детерминанти на клетката и действат като хормон-свързващи рецептори. Гликопротеините образуват адхезивни молекули, които карат клетките да се прикрепят една към друга, т.е. тесни междуклетъчни контакти.
клетка- това е не само течност, ензими и други вещества, но и високоорганизирани структури, наречени вътреклетъчни органели. Органелите за клетката са не по-малко важни от нейните химически компоненти. Така че, при липса на органели като митохондриите, доставката на енергия, извлечена от хранителни вещества, веднага ще намалее с 95%.
Повечето органели в клетката са покрити мембранисъставен предимно от липиди и протеини. Има клетъчни мембрани, ендоплазмен ретикулум, митохондрии, лизозоми, апарат на Голджи.
Липидиса неразтворими във вода, така че създават бариера в клетката, която предотвратява движението на вода и водоразтворими вещества от едно отделение в друго. Протеиновите молекули обаче правят мембраната пропусклива за различни веществачрез специализирани структури, наречени пори. Много други мембранни протеини са ензими, които катализират множество химична реакциякоито ще бъдат обсъдени в следващите глави.
Клетъчна (или плазмена) мембранае тънка, гъвкава и еластична структура с дебелина само 7,5-10 nm. Състои се главно от протеини и липиди. Приблизителното съотношение на неговите компоненти е както следва: протеини - 55%, фосфолипиди - 25%, холестерол - 13%, други липиди - 4%, въглехидрати - 3%.
липиден слой на клетъчната мембранапредотвратява проникването на вода. Основата на мембраната е липиден бислой - тънък липиден филм, състоящ се от два монослоя и напълно покриващ клетката. В цялата мембрана има протеини под формата на големи глобули.
Схематично представяне на клетъчната мембрана, отразяващо основните й елементи- фосфолипиден двуслой и голям бройпротеинови молекули, стърчащи над повърхността на мембраната.
Въглехидратните вериги са прикрепени към протеините на външната повърхност
и към допълнителни протеинови молекули вътре в клетката (това не е показано на фигурата).
липиден двуслойсе състои главно от фосфолипидни молекули. Единият край на такава молекула е хидрофилен, т.е. разтворим във вода (върху него е разположена фосфатна група), другият е хидрофобен, т.е. разтворим само в мазнини (съдържа мастна киселина).
Поради факта, че хидрофобната част на молекулата фосфолипидотблъсква водата, но се привлича от подобни части на едни и същи молекули, фосфолипидите имат естественото свойство да се прикрепят един към друг в дебелината на мембраната, както е показано на фиг. 2-3. Хидрофилната част с фосфатна група образува две мембранни повърхности: външната, която е в контакт с извънклетъчната течност, и вътрешната, която е в контакт с вътреклетъчната течност.
Среден липиден слойнепроницаем за йони и водни разтвориглюкоза и урея. Мастноразтворимите вещества, включително кислород, въглероден диоксид, алкохол, напротив, лесно проникват в тази област на мембраната.
молекулихолестеролът, който е част от мембраната, също са естествени липиди, тъй като тяхната стероидна група има висока разтворимост в мазнини. Тези молекули изглежда са разтворени в липидния двуслой. Основната им цел е регулирането на пропускливостта (или непроницаемостта) на мембраните за водоразтворимите компоненти на телесните течности. Освен това холестеролът е основният регулатор на вискозитета на мембраната.
Протеини на клетъчните мембрани. На фигурата се виждат глобуларни частици в липидния двуслой - това са мембранни протеини, повечето от които са гликопротеини. Има два вида мембранни протеини: (1) интегрални, които проникват през мембраната; (2) периферни, които стърчат само над едната повърхност, без да достигат до другата.
Много интегрални протеиниобразуват канали (или пори), през които водата и водоразтворимите вещества, особено йони, могат да дифундират във вътрешно- и извънклетъчната течност. Поради селективността на каналите някои вещества дифундират по-добре от други.
Други интегрални протеинифункционират като протеини носители, осъществяващи транспорта на вещества, за които липидният бислой е непроницаем. Понякога протеините носители действат в посока, противоположна на дифузията, такъв транспорт се нарича активен. Някои интегрални протеини са ензими.
Интегрални мембранни протеинимогат също да служат като рецептори за водоразтворими вещества, включително пептидни хормони, тъй като мембраната е непроницаема за тях. Взаимодействието на рецепторен протеин с определен лиганд води до конформационни промени в протеиновата молекула, което от своя страна стимулира ензимната активност на вътреклетъчния сегмент на протеиновата молекула или предаването на сигнал от рецептора в клетката с помощта на втори месинджър. Така интегралните протеини, вградени в клетъчната мембрана, я включват в процеса на пренос на информация за външната среда в клетката.
Молекули на периферни мембранни протеиничесто се свързва с интегрални протеини. Повечето периферни протеини са ензими или играят ролята на диспечер за транспортиране на вещества през мембранните пори.
Клетъчната мембрана има доста сложна структуракоето може да се види с електронен микроскоп. Грубо казано, той се състои от двоен слой липиди (мазнини), в който на различни места са включени различни пептиди (протеини). Общата дебелина на мембраната е около 5-10 nm.
Общ планструктурата на клетъчната мембрана е универсална за целия жив свят. Животинските мембрани обаче съдържат включвания на холестерол, което определя неговата твърдост. Разликата между мембраните на различните царства организми се отнася главно до надмембранните образувания (слоеве). Така че при растенията и гъбите над мембраната (отвън) има клетъчна стена. При растенията се състои главно от целулоза, а при гъбите - от веществото хитин. При животните епимембранният слой се нарича гликокаликс.
Друго име на клетъчната мембрана е цитоплазмена мембранаили плазмена мембрана.
По-задълбочено изследване на структурата на клетъчната мембрана разкрива много от нейните особености, свързани с изпълняваните функции.
Липидният двуслой се състои главно от фосфолипиди. Това са мазнини, единият край на които съдържа остатък от фосфорна киселина, който има хидрофилни свойства (тоест привлича водни молекули). Вторият край на фосфолипида е верига от мастни киселини, които имат хидрофобни свойства (не образуват водородни връзки с вода).
Фосфолипидните молекули в клетъчната мембрана се подреждат в два реда, така че хидрофобните им „краища“ са вътре, а хидрофилните „глави“ са отвън. Оказва се доста здрава структура, която предпазва съдържанието на клетката от външната среда.
Протеиновите включвания в клетъчната мембрана са неравномерно разпределени, освен това са подвижни (тъй като фосфолипидите в бислоя имат странична подвижност). От 70-те години на XX век хората започват да говорят за течно-мозаечна структура на клетъчната мембрана.
В зависимост от това как протеинът е част от мембраната, има три вида протеини: интегрални, полуинтегрални и периферни. Интегралните протеини преминават през цялата дебелина на мембраната, а краищата им стърчат от двете й страни. Те изпълняват основно транспортна функция. При полуинтегралните протеини единият край е разположен в дебелината на мембраната, а вторият излиза навън (отвън или отвътре). Те изпълняват ензимни и рецепторни функции. Периферните протеини се намират на външната или вътрешната повърхност на мембраната.
Структурните особености на клетъчната мембрана показват, че тя е основният компонент на повърхностния комплекс на клетката, но не и единственият. Другите му компоненти са надмембранният слой и подмембранният слой.
Гликокаликсът (супрамембранен слой на животните) се образува от олигозахариди и полизахариди, както и от периферни протеини и изпъкнали части от интегрални протеини. Компонентите на гликокаликса изпълняват рецепторна функция.
Освен гликокаликса, животинските клетки имат и други надмембранни образувания: слуз, хитин, перилема (подобно на мембрана).
Супрамембранната формация при растенията и гъбите е клетъчната стена.
Подмембранният слой на клетката е повърхностната цитоплазма (хиалоплазма) с включена в нея поддържащо-контрактилна система на клетката, чиито фибрили взаимодействат с протеините, изграждащи клетъчната мембрана. Чрез такива съединения от молекули се предават различни сигнали.