Bildung von Seitenwurzeln. Die Wurzel, ihre Funktionen. Arten von Wurzeln und Wurzelsystemen
Die Wurzel ist das unterirdische Achsenelement der Pflanzen, ihr wichtigster Teil, ihr vegetatives Hauptorgan. Dank der Wurzel wird die Pflanze im Boden verankert und dort während ihres gesamten Lebenszyklus gehalten und wird auch mit Wasser, Mineralien und darin enthaltenen Nährstoffen versorgt. Es gibt verschiedene Arten und Arten von Wurzeln. Jeder von ihnen hat seine eigenen charakteristischen Merkmale. In diesem Artikel werden wir die bestehenden Arten von Wurzeln und Arten von Wurzelsystemen betrachten. Wir werden uns auch mit ihren charakteristischen Merkmalen vertraut machen.
Welche Arten von Wurzeln gibt es?
Die Standardwurzel zeichnet sich durch eine fadenförmige oder schmalzylindrische Form aus. In vielen Pflanzen werden neben der Hauptwurzel (Hauptwurzel) auch andere Arten von Wurzeln entwickelt - seitlich und zufällig. Schauen wir uns genauer an, was sie sind.
Hauptwurzel
Dieses Pflanzenorgan entwickelt sich aus der Keimwurzel des Samens. Es gibt immer eine Hauptwurzel (andere Arten von Pflanzenwurzeln sind normalerweise Plural). Es verbleibt während des gesamten Lebenszyklus in der Pflanze.
Die Wurzel zeichnet sich durch positiven Geotropismus aus, dh sie vertieft sich aufgrund der Schwerkraft vertikal nach unten in das Substrat.
zufällige Wurzeln
Adventiv bezeichnet die Arten von Pflanzenwurzeln, die an ihren anderen Organen gebildet werden. Diese Organe können Stängel, Blätter, Triebe usw. sein. Getreide hat beispielsweise sogenannte primäre Adventivwurzeln, die im Stängel des Samenkeims abgelegt werden. Sie entwickeln sich im Prozess der Samenkeimung fast gleichzeitig mit der Hauptwurzel.
Es gibt auch blattadventive Arten von Wurzeln (gebildet als Ergebnis der Verwurzelung von Blättern), Stängel oder Knoten (gebildet aus Rhizomen, oberirdischen oder unterirdischen Stammknoten) usw. An den unteren Knoten werden starke Wurzeln gebildet, die als bezeichnet werden Antenne (oder tragend).
Das Auftreten von Adventivwurzeln bestimmt die Fähigkeit der Pflanze zur vegetativen Vermehrung.
Seitliche Wurzeln
Seitenwurzeln werden als Seitenwurzeln bezeichnet, die als Seitenast entstehen. Sie können sich sowohl an der Haupt- als auch an der Nebenwurzel bilden. Außerdem können sie von den seitlichen abzweigen, wodurch seitliche Wurzeln höherer Ordnung (erster, zweiter und dritter) gebildet werden.
Große Seitenorgane sind durch Quergeotropismus gekennzeichnet, dh ihr Wachstum erfolgt in nahezu horizontaler Position oder in einem Winkel zur Bodenoberfläche.
Was ist das Wurzelsystem?
Als Wurzelsystem werden alle Arten und Arten von Wurzeln bezeichnet, die eine Pflanze hat (dh ihre Gesamtheit). Je nach Wuchsverhältnis der Haupt-, Seiten- und Nebenwurzeln werden Art und Charakter bestimmt.
Arten von Wurzelsystemen
Wenn die Hauptwurzel sehr gut entwickelt und unter den Wurzeln einer anderen Art auffällt, bedeutet dies, dass die Pflanze ein Stabsystem hat. Es kommt hauptsächlich in zweikeimblättrigen Pflanzen vor.
Das Wurzelsystem dieses Typs zeichnet sich durch eine tiefe Keimung in den Boden aus. So können zum Beispiel die Wurzeln einiger Gräser bis zu einer Tiefe von 10-12 Metern vordringen (Distel, Luzerne). Die Eindringtiefe von Baumwurzeln kann in einigen Fällen 20 m erreichen.
Wenn Adventivwurzeln stärker ausgeprägt sind und sich in großer Zahl entwickeln und die Hauptwurzel durch langsames Wachstum gekennzeichnet ist, wird ein Wurzelsystem gebildet, das als faserig bezeichnet wird.

In der Regel zeichnet sich auch ein Teil der krautigen Pflanzen durch ein solches System aus. Obwohl die Wurzeln des Fasersystems nicht so tief eindringen wie die des Stabsystems, umflechten sie die angrenzenden Bodenpartikel besser. Viele lose Strauch- und Rhizom-Gräser, die reichlich faserige, dünne Wurzeln bilden, werden häufig verwendet, um Schluchten, Böden an Hängen usw. zu reparieren.

modifizierte Wurzeln
Neben den oben beschriebenen typischen gibt es noch andere Arten von Wurzeln und Wurzelsystemen. Sie werden modifiziert genannt.
Speicherwurzeln
Die Bestände umfassen Hackfrüchte und Wurzelknollen.
Eine Hackfrucht ist eine Verdickung der Hauptwurzel aufgrund der Ablagerung von Nährstoffen darin. Auch der untere Teil des Stängels ist an der Bildung der Wurzelfrucht beteiligt. Besteht hauptsächlich aus Speicherbasisgewebe. Beispiele für Hackfrüchte sind Petersilie, Radieschen, Karotten, Rüben usw.

Handelt es sich bei den verdickten Speicherwurzeln um Seiten- und Adventivwurzeln, so spricht man von Wurzelknollen (Zapfen). Sie werden in Kartoffeln, Süßkartoffeln, Dahlien usw. entwickelt.
Luftwurzeln
Dies sind Seitenwurzeln, die im oberirdischen Teil wachsen. In einer Zahl vorhanden tropische Pflanzen. Wasser und Sauerstoff werden aus der Luft aufgenommen. Verfügbar in tropischen Pflanzen, die unter Mangelbedingungen wachsen Mineralien.

Atmungswurzeln
Dies ist eine Art seitliche Wurzeln, die nach oben wachsen und sich über die Oberfläche des Substrats, Wasser, erheben. Solche Wurzeltypen werden in Pflanzen gebildet, die auf zu feuchten Böden unter Sumpfbedingungen wachsen. Mit Hilfe solcher Wurzeln erhält die Vegetation den fehlenden Sauerstoff aus der Luft.
Stützende (brettförmige) Wurzeln
Diese Arten von Baumwurzeln sind charakteristisch für große Arten (Buche, Ulme, Pappel, Tropenholz usw.) Es handelt sich um dreieckige vertikale Auswüchse, die von Seitenwurzeln gebildet werden und nahe oder über der Bodenoberfläche verlaufen. Sie werden auch brettförmig genannt, weil sie Brettern ähneln, die an einen Baum gelehnt sind.
Saugwurzeln (Haustoria)
Dies ist eine Art zusätzliche Adventivwurzel, die sich am Stängel von Kletterpflanzen entwickelt. Mit ihrer Hilfe haben Pflanzen die Fähigkeit, sich an eine bestimmte Stütze zu heften und hochzuklettern (weben). Solche Wurzeln gibt es zum Beispiel in zähem Ficus, Efeu etc.

Einziehbare (kontraktile) Wurzeln
Charakteristisch für Pflanzen, deren Wurzel an der Basis in Längsrichtung stark reduziert ist. Ein Beispiel wären Pflanzen mit Zwiebeln. Einziehbare Wurzeln bieten Zwiebeln und Hackfrüchten eine gewisse Aussparung im Boden. Darüber hinaus bestimmt ihre Anwesenheit den festen Sitz von Rosetten (z. B. in einem Löwenzahn) am Boden sowie die unterirdische Position des vertikalen Rhizoms und des Wurzelhalses.
Mykorrhiza (Pilzwurzel)
Mykorrhiza ist eine Symbiose (für beide Seiten vorteilhaftes Zusammenleben) der Wurzeln höherer Pflanzen mit Pilzhyphen, die sie als Wurzelhaare flechten. Pilze versorgen Pflanzen mit Wasser und darin gelösten Nährstoffen. Pflanzen wiederum versorgen Pilze mit organischen Stoffen, die sie für ihre Lebenstätigkeit benötigen.
Mykorrhiza ist in den Wurzeln vieler höherer Pflanzen, insbesondere holziger, inhärent.
bakterielle Knötchen
Dies sind modifizierte Seitenwurzeln, die für eine symbiotische Kohabitation mit stickstofffixierenden Bakterien angepasst sind. Die Bildung von Knötchen erfolgt durch das Eindringen junger Wurzeln in das Innere. Ein solches für beide Seiten vorteilhaftes Zusammenleben ermöglicht es Pflanzen, Stickstoff aufzunehmen, den Bakterien aus der Luft in eine für sie zugängliche Form übertragen. Bakterien hingegen erhalten einen speziellen Lebensraum, in dem sie funktionieren können, ohne mit anderen Bakterienarten zu konkurrieren. Darüber hinaus verwenden sie Substanzen, die in den Wurzeln der Vegetation vorhanden sind.

Bakterienknollen sind typisch für Pflanzen aus der Familie der Leguminosen, die als Meliorationsmittel in Fruchtfolgen weit verbreitet sind, um Böden mit Stickstoff anzureichern. Pfahlwurzelleguminosen wie blaue und gelbe Luzerne, rote und Esparsette, Hornheuschrecke usw. gelten als die besten stickstofffixierenden Pflanzen.
Zusätzlich zu den oben genannten Metamorphosen gibt es andere Arten von Wurzeln, wie z. B. Stützwurzeln (helfen, den Stamm zu stärken), Stelzwurzeln (helfen Pflanzen, nicht in flüssigen Schlamm einzusinken) und Wurzelausläufer (haben zufällige Knospen und sorgen für vegetative Vermehrung).
Die Wurzel entstand phylogenetisch später als Stängel und Blatt – im Zusammenhang mit dem Übergang der Pflanzen zum Leben an Land und entstand wahrscheinlich aus wurzelartigen unterirdischen Ästen. Die Wurzel hat weder Blätter noch Knospen, die in einer bestimmten Reihenfolge angeordnet sind. Es zeichnet sich durch apikales Längenwachstum aus, seine Seitenäste entstehen aus inneren Geweben, der Wachstumspunkt ist mit einer Wurzelkappe bedeckt. Das Wurzelsystem wird während des gesamten Lebens des Pflanzenorganismus gebildet. Manchmal kann die Wurzel als Ablagerungsort bei der Nährstoffversorgung dienen. In diesem Fall wird es modifiziert.
Root-Typen
Die Hauptwurzel wird während der Samenkeimung aus der Keimwurzel gebildet. Es hat seitliche Wurzeln.
Adventivwurzeln entwickeln sich an Stängeln und Blättern.
Seitenwurzeln sind Äste beliebiger Wurzeln.
Jede Wurzel (Haupt-, Seiten-, Nebenwurzel) kann sich verzweigen, wodurch die Oberfläche des Wurzelsystems erheblich vergrößert wird, was zu einer besseren Stärkung der Pflanze im Boden und einer verbesserten Ernährung beiträgt.
Arten von Wurzelsystemen
Es gibt zwei Haupttypen von Wurzelsystemen: Pfahlwurzeln, die eine gut entwickelte Hauptwurzel haben, und faserige. Das Faserwurzelsystem besteht aus einer großen Anzahl von Adventivwurzeln gleicher Größe. Die gesamte Wurzelmasse besteht aus Seiten- oder Adventivwurzeln und sieht aus wie ein Lappen.
Ein stark verzweigtes Wurzelsystem bildet eine riesige Aufnahmefläche. Zum Beispiel,
- die Gesamtlänge der Winterroggenwurzeln erreicht 600 km;
- Länge der Wurzelhaare - 10.000 km;
- Die Gesamtfläche der Wurzeln beträgt 200 m 2.
Diese ist um ein Vielfaches größer als die Fläche der oberirdischen Masse.

Wenn die Pflanze eine gut definierte Hauptwurzel hat und sich Adventivwurzeln entwickeln, dann wird ein gemischtartiges Wurzelsystem (Kohl, Tomate) gebildet.
Äußere Struktur der Wurzel. Die innere Struktur der Wurzel
Wurzelzonen

Wurzelkappe
Die Wurzel wächst mit ihrer Spitze, wo sich die jungen Zellen des Bildungsgewebes befinden, in die Länge. Der wachsende Teil ist mit einer Wurzelkappe bedeckt, die die Wurzelspitze vor Beschädigungen schützt und die Bewegung der Wurzel im Boden während des Wachstums erleichtert. Letztere Funktion wird aufgrund der Eigenschaft der Außenwände der Wurzelkappe erfüllt, mit Schleim bedeckt zu sein, was die Reibung zwischen der Wurzel und den Erdpartikeln verringert. Sie können sogar Bodenpartikel auseinanderdrücken. Die Zellen der Wurzelkappe leben und enthalten oft Stärkekörner. Die Zellen der Kappe werden aufgrund der Teilung ständig aktualisiert. Beteiligt sich an positiven geotropischen Reaktionen (Richtung des Wurzelwachstums zum Erdmittelpunkt).

Die Zellen der Teilungszone teilen sich aktiv, die Länge dieser Zone variiert bei verschiedenen Arten und bei verschiedenen Wurzeln derselben Pflanze.
Hinter der Teilungszone befindet sich eine Erweiterungszone (Wachstumszone). Die Länge dieser Zone überschreitet einige Millimeter nicht.
Wenn das lineare Wachstum abgeschlossen ist, beginnt die dritte Stufe der Wurzelbildung - ihre Differenzierung, eine Zone der Differenzierung und Spezialisierung von Zellen (oder eine Zone von Wurzelhaaren und Absorption) wird gebildet. In dieser Zone sind bereits die äußere Schicht des Epiblemas (Rhizoderm) mit Wurzelhaaren, die Schicht der primären Rinde und der zentrale Zylinder zu unterscheiden.
Die Struktur des Wurzelhaars
Wurzelhaare sind stark verlängerte Auswüchse der äußeren Zellen, die die Wurzel bedecken. Die Anzahl der Wurzelhaare ist sehr hoch (von 200 bis 300 Haare pro 1 mm2). Ihre Länge erreicht 10 mm. Haare bilden sich sehr schnell (bei jungen Sämlingen eines Apfelbaums in 30-40 Stunden). Wurzelhaare sind kurzlebig. Sie sterben in 10-20 Tagen ab und neue wachsen auf dem jungen Teil der Wurzel. Dadurch wird die Entwicklung neuer Bodenhorizonte durch die Wurzel gewährleistet. Die Wurzel wächst kontinuierlich und bildet immer neue Bereiche von Wurzelhaaren. Haare können nicht nur fertige Stofflösungen aufnehmen, sondern auch zur Auflösung bestimmter Bodenstoffe beitragen und diese dann aufnehmen. Der Bereich der Wurzel, wo die Wurzelhaare abgestorben sind, kann noch einige Zeit Wasser aufnehmen, wird dann aber mit Kork überzogen und verliert diese Fähigkeit.
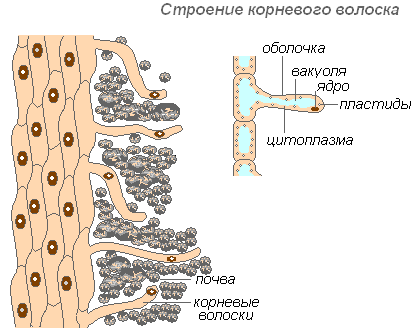
Die Haarhülle ist sehr dünn, was die Aufnahme von Nährstoffen erleichtert. Fast die gesamte Haarzelle ist von einer Vakuole besetzt, die von einer dünnen Zytoplasmaschicht umgeben ist. Der Zellkern befindet sich an der Spitze der Zelle. Um die Zelle herum bildet sich eine Schleimhaut, die das Verkleben der Wurzelhaare mit Bodenpartikeln fördert, was ihren Kontakt verbessert und die Hydrophilie des Systems erhöht. Die Absorption wird durch die Sekretion von Säuren (Kohlensäure, Äpfelsäure, Zitronensäure) durch Wurzelhaare erleichtert, die Mineralsalze lösen.
Wurzelhaare spielen auch eine mechanische Rolle - sie dienen als Stütze für die Wurzelspitze, die zwischen den Bodenpartikeln hindurchgeht.
Unter einem Mikroskop ist auf einem Querschnitt der Wurzel in der Absorptionszone ihre Struktur auf der Zell- und sichtbar Gewebeebenen. An der Oberfläche der Wurzel befindet sich das Rhizoderm, darunter die Rinde. Die äußere Schicht der Rinde ist das Exoderm, nach innen davon das Hauptparenchym. Seine dünnwandigen lebenden Zellen erfüllen eine Speicherfunktion, leiten Nährlösungen in radialer Richtung - vom absorbierenden Gewebe zu den Gefäßen des Holzes. Sie synthetisieren auch eine Reihe von lebenswichtigen organischen Substanzen für die Pflanze. Die innere Schicht der Rinde ist das Endoderm. Nährlösungen, die von der Rinde zum zentralen Zylinder durch die Zellen des Endoderms gelangen, passieren nur den Protoplasten der Zellen.

Die Rinde umgibt den zentralen Zylinder der Wurzel. Sie grenzt an eine Zellschicht, die ihre Teilungsfähigkeit lange behält. Das ist das Perizykel. Perizyklische Zellen führen zu Seitenwurzeln, Adnexknospen und sekundären Bildungsgeweben. Vom Perizykel nach innen, in der Mitte der Wurzel, befinden sich leitfähige Gewebe: Bast und Holz. Zusammen bilden sie einen radialen Leitstrahl.
Das Leitungssystem der Wurzel leitet Wasser und Mineralien von der Wurzel zum Stamm (Aufwärtsströmung) und organische Stoffe vom Stamm zur Wurzel (Abwärtsströmung). Es besteht aus vaskulären Faserbündeln. Die Hauptbestandteile des Bündels sind Abschnitte des Phloems (durch das sich Substanzen zur Wurzel bewegen) und des Xylems (durch das sich Substanzen von der Wurzel bewegen). Die Hauptleitungselemente des Phloems sind Siebröhren, Xyleme sind Tracheen (Gefäße) und Tracheiden.
Wurzellebensprozesse
Wassertransport an der Wurzel
Aufnahme von Wasser durch Wurzelhaare aus der Bodennährlösung und dessen Weiterleitung in radialer Richtung entlang der Zellen der Primärrinde durch die Durchgangszellen in der Endodermis zum Xylem des radialen Leitbündels. Die Intensität der Wasseraufnahme durch die Wurzelhaare wird Saugkraft (S) genannt, sie ist gleich der Differenz zwischen osmotischem (P) und Turgordruck (T): S=P-T.
Wenn der osmotische Druck gleich dem Turgordruck (P = T) ist, dann S = 0, hört das Wasser auf, in die Wurzelhaarzelle zu fließen. Wenn die Konzentration der Substanzen in der Bodennährlösung höher ist als in der Zelle, verlässt Wasser die Zellen und es kommt zur Plasmolyse - die Pflanzen verdorren. Dieses Phänomen wird bei trockenem Boden sowie bei übermäßigem Einsatz von Mineraldünger beobachtet. Innerhalb der Wurzelzellen nimmt die Saugkraft der Wurzel vom Rhizoderm zum zentralen Zylinder zu, sodass sich Wasser entlang des Konzentrationsgradienten bewegt (d. h. von einem Ort mit höherer Konzentration zu einem Ort mit niedrigerer Konzentration) und einen Wurzeldruck erzeugt das eine Wassersäule entlang der Xylemgefäße hebt und eine aufwärts gerichtete Strömung bildet. Es kann auf blattlosen Frühlingsstämmen gefunden werden, wenn "Saft" geerntet wird, oder auf geschnittenen Stümpfen. Das Abfließen von Wasser aus Holz, frischen Stümpfen und Blättern wird als "Weinen" von Pflanzen bezeichnet. Wenn die Blätter blühen, erzeugen sie auch eine Saugkraft und ziehen Wasser an sich - in jedem Gefäß bildet sich eine durchgehende Wassersäule - Kapillarspannung. Der Wurzeldruck ist der untere Motor der Wasserströmung, die Saugkraft der Blätter der obere. Sie können dies mit Hilfe einfacher Experimente bestätigen.
Aufnahme von Wasser durch Wurzeln
Ziel: Finden Sie die Hauptfunktion der Wurzel heraus.
Was wir tun: B. eine auf nassem Sägemehl gewachsene Pflanze, schütteln Sie ihr Wurzelsystem ab und senken Sie ihre Wurzeln in ein Glas Wasser. Gießen Sie eine dünne Schicht über das Wasser, um es vor Verdunstung zu schützen. Pflanzenöl und notieren Sie das Niveau.

Was wir beobachten: Nach ein oder zwei Tagen fiel das Wasser im Tank unter die Marke.

Ergebnis: Daher saugten die Wurzeln das Wasser an und brachten es zu den Blättern.
Ein weiteres Experiment kann durchgeführt werden, um die Aufnahme von Nährstoffen durch die Wurzel zu beweisen.
Was wir tun: Wir schneiden den Stängel der Pflanze ab und hinterlassen einen 2-3 cm hohen Stumpf.Wir legen einen 3 cm langen Gummischlauch auf den Stumpf und setzen ein gebogenes Glasrohr mit einer Höhe von 20-25 cm auf das obere Ende.
Was wir beobachten: das Wasser im Glasrohr steigt und fließt heraus.
Ergebnis: dies beweist, dass die Wurzel Wasser aus dem Boden in den Stängel aufnimmt.

Beeinflusst die Wassertemperatur die Wasseraufnahme durch die Wurzel?
Ziel: Finden Sie heraus, wie sich die Temperatur auf den Root-Betrieb auswirkt.
Was wir tun: Ein Glas sollte mit warmem Wasser (+17-18ºС) und das andere mit kaltem Wasser (+1-2ºС) gefüllt sein.
Was wir beobachten: im ersten Fall wird Wasser reichlich freigesetzt, im zweiten - wenig oder hört ganz auf.
Ergebnis: dies ist ein Beweis dafür, dass die Temperatur einen starken Einfluss auf die Wurzelleistung hat.

Warmes Wasser wird aktiv von den Wurzeln aufgenommen. Der Wurzeldruck steigt.

Kaltes Wasser wird von den Wurzeln schlecht aufgenommen. In diesem Fall sinkt der Wurzeldruck.
mineralische Ernährung
Die physiologische Rolle der Mineralien ist sehr groß. Sie sind die Basis für die Synthese organische Verbindungen, sowie Faktoren, die den Aggregatzustand von Kolloiden verändern, d.h. direkt den Stoffwechsel und die Struktur des Protoplasten beeinflussen; wirken als Katalysatoren für biochemische Reaktionen; beeinflussen den Turgor der Zelle und die Permeabilität des Protoplasmas; sind die Zentren elektrischer und radioaktiver Phänomene in Pflanzenorganismen.
Es wurde festgestellt, dass die normale Entwicklung von Pflanzen nur in Gegenwart von drei Nichtmetallen in der Nährlösung - Stickstoff, Phosphor und Schwefel - und vier Metallen - Kalium, Magnesium, Kalzium und Eisen - möglich ist. Jedes dieser Elemente hat einen individuellen Wert und kann nicht durch ein anderes ersetzt werden. Dies sind Makronährstoffe, ihre Konzentration in der Pflanze beträgt 10 -2 -10%. Für die normale Entwicklung von Pflanzen werden Mikroelemente benötigt, deren Konzentration in der Zelle 10 -5 -10 -3% beträgt. Dies sind Bor, Kobalt, Kupfer, Zink, Mangan, Molybdän usw. Alle diese Elemente kommen im Boden vor, aber manchmal in unzureichenden Mengen. Daher werden mineralische und organische Düngemittel auf den Boden aufgebracht.
Die Pflanze wächst und entwickelt sich normal, wenn die Umgebung der Wurzeln alle notwendigen Nährstoffe enthält. Der Boden ist eine solche Umgebung für die meisten Pflanzen.
Atem der Wurzel
Für ein normales Wachstum und eine normale Entwicklung der Pflanze ist es notwendig, dass frische Luft in die Wurzel gelangt. Lassen Sie uns überprüfen, ob es ist?
Ziel: Brauchen Wurzeln Luft?
Was wir tun: Nehmen wir zwei identische Gefäße mit Wasser. Wir setzen in jedes Gefäß sich entwickelnde Sämlinge. Das Wasser in einem der Gefäße sättigen wir jeden Tag mit einer Sprühflasche mit Luft. Gießen Sie auf die Wasseroberfläche im zweiten Gefäß eine dünne Schicht Pflanzenöl, da dies den Luftstrom in das Wasser verzögert.

Was wir beobachten: Nach einer Weile hört die Pflanze im zweiten Gefäß auf zu wachsen, verdorrt und stirbt schließlich ab.

Ergebnis: Der Tod der Pflanze tritt aufgrund des Luftmangels auf, der für die Atmung der Wurzel erforderlich ist.
Root-Modifikationen
Bei manchen Pflanzen werden Reservenährstoffe in den Wurzeln abgelagert. Sie reichern Kohlenhydrate, Mineralsalze, Vitamine und andere Substanzen an. Solche Wurzeln werden stark dicker und erhalten ein ungewöhnliches Aussehen. Sowohl die Wurzel als auch der Stamm sind an der Bildung von Hackfrüchten beteiligt.
Wurzeln
Sammeln sich Reservestoffe in der Hauptwurzel und an der Stängelbasis des Hauptsprosses, bilden sich Hackfrüchte (Karotten). Wurzelbildende Pflanzen sind meist zweijährig. Im ersten Lebensjahr blühen sie nicht und reichern viele Nährstoffe in Hackfrüchten an. Auf der zweiten blühen sie schnell, nutzen die angesammelten Nährstoffe und bilden Früchte und Samen.
Wurzelknollen

Bei Dahlien reichern sich Reservestoffe in Adventivwurzeln an und bilden Wurzelknollen.
bakterielle Knötchen
Die Seitenwurzeln von Klee, Lupine, Luzerne sind eigentümlich verändert. Bakterien siedeln sich in jungen Seitenwurzeln an, was zur Aufnahme von gasförmigem Stickstoff aus der Bodenluft beiträgt. Solche Wurzeln nehmen die Form von Knötchen an. Dank dieser Bakterien sind diese Pflanzen in der Lage, auf stickstoffarmen Böden zu leben und diese fruchtbarer zu machen.
gestelzt

Eine in der Gezeitenzone wachsende Rampe entwickelt Stelzwurzeln. Hoch über dem Wasser halten sie große Blatttriebe auf instabilem, schlammigem Boden.
Luft

Tropische Pflanzen, die auf Ästen leben, entwickeln Luftwurzeln. Sie werden oft in Orchideen, Bromelien und einigen Farnen gefunden. Luftwurzeln hängen frei in der Luft, erreichen den Boden nicht und nehmen Feuchtigkeit von Regen oder Tau auf, der auf sie fällt.
Retraktoren

Bei Zwiebel- und Knollengewächsen, beispielsweise Krokussen, gibt es neben den zahlreichen Fadenwurzeln auch mehrere dickere, sogenannte Rückzugswurzeln. Reduzierend ziehen solche Wurzeln die Knolle tiefer in den Boden.
Säulenförmig

Ficus entwickeln säulenförmige oberirdische Wurzeln oder Stützwurzeln.
Boden als Lebensraum für Wurzeln
Der Boden für Pflanzen ist die Umgebung, aus der sie Wasser und Nährstoffe erhält. Der Gehalt an Mineralien im Boden hängt von den Besonderheiten des Ausgangsgesteins, der Aktivität von Organismen, der Vitalaktivität der Pflanzen selbst und der Art des Bodens ab.
Bodenpartikel konkurrieren mit den Wurzeln um Feuchtigkeit und halten sie auf ihrer Oberfläche. Dies ist das sogenannte gebundene Wasser, das in hygroskopisch und filmartig unterteilt wird. Es wird durch die Kräfte der molekularen Anziehung gehalten. Die der Pflanze zur Verfügung stehende Feuchtigkeit wird durch Kapillarwasser repräsentiert, das sich in den kleinen Poren des Bodens konzentriert.
Es entstehen antagonistische Beziehungen zwischen der Feuchtigkeits- und der Luftphase des Bodens. Je mehr große Poren im Boden sind, desto besser ist das Gasregime dieser Böden, desto weniger Feuchtigkeit speichert der Boden. Das günstigste Wasser-Luft-Regime wird in Strukturböden aufrechterhalten, in denen sich Wasser und Luft gleichzeitig befinden und sich nicht gegenseitig stören - Wasser füllt die Kapillaren innerhalb der Strukturaggregate und Luft füllt die großen Poren zwischen ihnen.
Die Art der Wechselwirkung zwischen der Pflanze und dem Boden hängt weitgehend mit der Aufnahmefähigkeit des Bodens zusammen - der Fähigkeit, chemische Verbindungen zurückzuhalten oder zu binden.
Die Bodenmikroflora zersetzt organisches Material zu mehr einfache Verbindungen, beteiligt sich an der Bildung der Bodenstruktur. Die Art dieser Prozesse hängt von der Art des Bodens, der chemischen Zusammensetzung von Pflanzenresten, den physiologischen Eigenschaften von Mikroorganismen und anderen Faktoren ab. Bodentiere sind an der Bildung der Bodenstruktur beteiligt: Anneliden, Insektenlarven usw.
Als Ergebnis einer Kombination aus biologischen u Chemische Prozesse Im Boden bildet sich ein komplexer Komplex organischer Substanzen, der unter dem Begriff „Humus“ zusammengefasst wird.
Methode der Wasserkultur
Welche Salze eine Pflanze braucht und wie sie sich auf ihr Wachstum und ihre Entwicklung auswirken, wurde durch Experimente mit Wasserkulturen ermittelt. Die aquatische Kulturmethode ist die Kultivierung von Pflanzen nicht in Erde, sondern in einer wässrigen Lösung von Mineralsalzen. Je nach Ziel des Experiments können Sie ein separates Salz aus der Lösung ausschließen, seinen Gehalt verringern oder erhöhen. Es wurde festgestellt, dass stickstoffhaltige Düngemittel zum Wachstum von Pflanzen beitragen, phosphorhaltige Düngemittel die früheste Reifung von Früchten und kaliumhaltige Düngemittel den schnellsten Abfluss organischer Stoffe von den Blättern zu den Wurzeln. In diesem Zusammenhang wird empfohlen, stickstoffhaltige Düngemittel vor der Aussaat oder in der ersten Sommerhälfte auszubringen, die Phosphor und Kalium enthalten - in der zweiten Sommerhälfte.
Mit der Methode der Wasserkulturen konnte nicht nur der Bedarf einer Pflanze an Makroelementen festgestellt, sondern auch die Rolle verschiedener Mikroelemente ermittelt werden.
Derzeit gibt es Fälle, in denen Pflanzen mit Hydroponik- und Aeroponik-Methoden angebaut werden.
Hydroponik ist die Kultivierung von Pflanzen in mit Kies gefüllten Töpfen. Nährlösung enthalten notwendige Elemente, wird von unten in die Gefäße eingespeist.
Aeroponik ist die Luftkultur von Pflanzen. Bei dieser Methode befindet sich das Wurzelsystem in der Luft und wird automatisch (mehrmals innerhalb einer Stunde) mit einer schwachen Nährsalzlösung besprüht.
ORGANOGRAPHIE
Organographie - Beschreibung der Organe eines lebenden Organismus.
Organ- ein morphologisch differenzierter und funktionell spezialisierter Körperteil. In Übereinstimmung mit den zwei Hauptfunktionen: das Leben einer gegebenen Pflanze zu erhalten und sie in einer Anzahl nachfolgender Generationen zu reproduzieren, werden vegetative und generative (reproduktive) Organe unterschieden.
Zunächst untersuchen wir die vegetativen Organe, die das Sprosssystem und das Wurzelsystem bilden. Das Element des Wurzelsystems ist die Wurzel.
Wurzel
Die Wurzel entwickelte sich später als der Spross und stammt aus den Rhizoiden der ersten Landpflanzen - Rhinophyten.
Verschiedene Teile der Wurzel erfüllen unterschiedliche Funktionen und zeichnen sich durch bestimmte aus morphologische Merkmale. Diese Teile werden Zonen genannt. Zonen einer jungen Wurzel: Teilungszone, Wachstums- oder Dehnungszone, Absorptions- oder Absorptionszone und Leitungszone.
Außen ist das apikale Meristem von einer Wurzelkappe bedeckt. Lebende Parenchymzellen der Kappe produzieren Schleim. Die Schleimsubstanz ist ein stark hydratisiertes Pektinpolysaccharid. Es reichert sich in Vesikeln von Dictyosomen an, die es beim Verschmelzen mit der Plasmamembran in den Raum zwischen Plasmamembran und Zellmembran freisetzen. Dann gelangt der Schleim an die Oberfläche der Zellmembran, wo er kleine Tropfen bildet.
Durch diesen Schleimbelag bleiben Erdpartikel an der Wurzelspitze und den Wurzelhaaren haften. Der Schleim dient als absorbierende Oberfläche für den Ionenaustausch. Die Schleimhaut schützt die Wurzelspitze vor Schadstoffe Boden und vor Austrocknung, schafft günstige Bedingungen für die Ansiedlung nützlicher Bakterien.
Wenn die Wurzel wächst und die Wurzelkappe nach vorne drückt, werden die Zellen an der Peripherie der Kappe abgestoßen. Die Lebensdauer der Zellen vom Auftreten bis zur Abschuppung beträgt 4-9 Tage, abhängig von der Länge des Hutes und der Pflanzenart.
Die Root-Kappe steuert die Root-Georezeption. Der Ort der Wahrnehmung der Schwerkraft in der Kappe ist die zentrale Zellsäule - Columella, in der Amyloplasten als Statolithen (Schwerkraftrezeptoren) fungieren.
Das apikale Meristem besteht aus kleinen (10–20 µm Durchmesser) polyedrischen Zellen mit dichtem Zytoplasma und großen Kernen.
Sporenpflanzen haben eine monoinitiale Wurzelspitze.
In den Wurzeln von Samenpflanzen wurden zwei Haupttypen der apikalen Organisation beschrieben. In einem von ihnen haben alle Bereiche gemeinsame Initialen. Dies ist eine offene Art von apikalem Meristem, die wahrscheinlich phylogenetisch primitiv ist. Beim zweiten Typ ist jeder der drei Teile, d.h. Der zentrale (oder leitende) Zylinder, die primäre Rinde und die Wurzelkappe haben ihre eigenen Initialen. Dies ist eine geschlossene Art von apikalem Meristem.
Die Initialen, die für die Organisation des Zellwachstums in den ersten Stadien verantwortlich sind, verlieren im Allgemeinen ihre mitotische Aktivität in den nachfolgenden Stadien des Wurzelwachstums. Stattdessen zeigen Zellen, die etwas tiefer im Wurzelkörper liegen, mitotische Aktivität. Diese Beobachtungen führten zur Bildung der Theorie des Ruhezentrums im apikalen Meristem. Nach dieser Theorie teilen sich die distalsten Zellen (der distalste Teil des Organs, der am weitesten vom Mittelpunkt entfernt ist: das untere Ende der Wurzel) im Wurzelkörper (die ersten Initialen des Pleroms und des Periblema) selten, haben ungefähr das Gleiche Größe und haben niedrige NA- und Proteinsyntheseraten. Das Ruhezentrum hat eine halbkugel- oder scheibenförmige Form und variiert je nach Größe der Wurzel im Volumen. Die Zellen des Ruhezentrums teilen sich etwa 10-mal langsamer als benachbarte Zellen.
Bei experimentell geschädigten Wurzeln ist das ruhende Zentrum in der Lage, das Meristem wiederherzustellen. Das heißt, das Ruhezentrum spielt eine wesentliche Rolle bei der Bildung der Struktur und bei der Entwicklung der Wurzel.
Ein Merkmal der Wurzelspitze ist die Anordnung von Längsreihen von Zellen, die ihren Ursprung im apikalen Meristem haben.
Schichten des apikalen Meristems der Wurzel:
Dermatogen(Griechisch Derma - Haut, Genos - Geburt) - die äußere Schicht des apikalen Meristems, differenziert sich in das Rhizoderm. Dermatogenzellen der richtigen Form.
Pleroma(griechisches Pleroma - Füllung) - der zentrale Teil des apikalen Meristems differenziert sich in Zellen des zentralen Zylinders. Die Zellen sind entlang der Achse verlängert.
Gefahr(griechisches Periblema - Deckblatt) - eine Schicht des apikalen Meristems zwischen dem Dermatogen und dem Plerom, differenziert sich in Zellen des primären Kortex.
Der primäre Cortex differenziert sich auf der Ebene der Elongationszone. In der Kolonisationszone macht der primäre Kortex den größten Teil des Querschnitts aus. Es wird aufgrund seines Ursprungs aus dem primären Meristem "primär" genannt, aufgrund seiner peripheren Lage "Rinde".
Die äußeren Zellen der primären Rinde, die sich unter dem Rhizoderm befinden, werden Exoderm genannt. Die innere Schicht der primären Rinde ist das Endoderm.
Der Großteil der primären Rinde ist das Mesoderm. Das Mesoderm wird von locker liegenden Parenchymzellen gebildet, was zur Belüftung der Wurzelzellen beiträgt.
Die Wurzeln der Gymnospermen und der meisten Dikotylen werfen ihre Primärrinde früh ab. In ihnen bleiben die Zellen der primären Rinde parenchymal. Im Gegensatz dazu bleibt der primäre Cortex von Monokotylen während des gesamten Lebens der Wurzel bestehen, und seine Zellen bilden sekundäre verholzte Hüllen.
Das Zentrum der Wurzel wird von einem axialen Zylinder eingenommen - einer Stele. Beginnt sich in der Wachstumszone zu differenzieren. Die Bildung des Axialzylinders beginnt mit der Bildung seiner äußeren Schicht - Perizykel. Das Pericyclus gehört zu den primären lateralen Meristemen, seine Zellen behalten ihre meristematische Aktivität lange bei. Normalerweise wird der Pericyclus durch eine einzelne Zellschicht dargestellt, obwohl er auch mehrschichtig sein kann.
Bei den meisten Samenpflanzen werden seitliche Wurzeln in das Perizykel gelegt, die Rudimente von Adnexknospen erscheinen. Bei Arten mit sekundärem Wachstum ist das Pericyclus an der Bildung des Kambiums beteiligt und führt zur ersten Schicht des Phellogens.
Unter dem Perizykel werden prokambiale Zellen abgelegt, aus denen das primäre Phloem, später das primäre Xylem entsteht. Die Anzahl der Xylemrochen variiert bei verschiedenen Arten von zwei, drei, vier bis zu mehreren. Entsprechend der Anzahl der Xylemstrahlen werden die Wurzeln Diarch, Triarch, Tetrarch, Polyarch genannt. Tetrarchenwurzeln gelten als primär. Monocots haben eine Polyarchstele.
Auf diese Weise, Gesamtplan Die Primärstruktur der Wurzel und die morphologische Differenzierung der Gewebe zeigen ein hohes Maß an Spezialisierung und Anpassung der Wurzel an die Funktionen der Aufnahme und Weiterleitung von Wasser und Mineralien.
Seitliche Wurzelbildung
Seitenwurzeln entstehen an der Peripherie des zentralen Zylinders in unterschiedlichen Abständen vom apikalen Meristem. Da Sie werden in tief liegende Gewebe der Wurzel gelegt, ihr Ursprung wird als endogen bezeichnet (aus dem Griechischen "geboren im Inneren"). Seitenwurzeln von Gymnospermen und Angiospermen werden in das Perizykel gelegt. Die Zellen des Perizykels, die an der Bildung von Seitenwurzeln beteiligt sind, sind der Wurzelbogen.
Auch mehrere Zellschichten der Wurzelanlagen können vom Endoderm gebildet werden.
Bei einer diarchischen Wurzel liegt eine Seitenwurzel zwischen Phloem und Xylem, bei einer triarchischen, tetrarchnischen Wurzel gegenüber dem Xylem, bei einer polyarchischen Wurzel von Monokotylen liegt sie dem Phloem gegenüber. Perizykelzellen teilen sich in verschiedene Richtungen. Als Ergebnis wird ein kleiner Wurzeltuberkel gebildet. Das Wurzelrudiment drückt mit zunehmender Größe auf das Entoderm. Aber die Schalen des Endoderms sind hart. Daher steht das Endoderm vor. Die Ausbuchtung des Entoderms wird als Wurzeltasche bezeichnet.
Die Anlage der Wurzel passiert die primäre Rinde und sondert dabei Enzyme ab, die die Zellen der Rinde zerstören. In den frühen Entwicklungsstadien bildet dieses Rudiment die Wurzelkappe, apikale und primäre Meristeme. Im Laufe der Zeit werden aufgrund der Differenzierung von Parenchymzellen in Elemente von Xylem und Phloem die zentralen Zylinder der Seiten- und Elternwurzeln miteinander verbunden.
Bei Monokotylen bleibt die primäre Wurzelstruktur lebenslang erhalten und sekundäres Gewebe entsteht nicht. Mit zunehmendem Alter können die Zellgrößen zunehmen, die Zellmembranen verdicken, aber der allgemeine "Plan" der Wurzelstruktur bleibt unverändert.
Bei Gymnospermen und Dikotylen treten sekundäre Veränderungen auf, und es wird eine sekundäre Wurzelstruktur gebildet. Die Bildung der Sekundärstruktur der Wurzel ist in erster Linie mit der Aktivität des Kambiums verbunden. Das Kambium wird durch die Teilung jener prokambialen Zellen gebildet, die zwischen dem primären Phloem und dem primären Xylem undifferenziert bleiben. Dann zeigen auch die Zellen des Perizykels, die außerhalb der Xylemvorsprünge lokalisiert sind, kambiale Aktivität.
Das Kambium lagert sekundäre Xylemzellen im Zentrum und sekundäre Phloemzellen an der Peripherie ab. So entstehen leitfähige Bündel. Sie werden so viel gebildet, wie es Strahlen des primären Xylems gab. Unter dem Druck wachsender Sekundärgewebe werden die Zellen des Primärphloems abgeflacht. Das primäre Xylem behält seine zentrale Position und ist als Sternchen sichtbar. Im Laufe der Zeit wird auch das primäre Xylem ausgelöscht.
In der Sekundärstruktur der Wurzel fungieren die Primärgewebe nicht als leitende Elemente. Der Stoffstrom in beide Richtungen erfolgt durch Sekundärgewebe.
Das aus dem Perizykel stammende Kambium bildet breite Parenchymstrahlen, die zwischen den offenen Kollateralbündeln liegen. Dies sind die primären Kernstrahlen. Sie stellen eine physiologische Verbindung zwischen dem zentralen Teil der Wurzel und der primären Rinde her. Später können schmalere sekundäre Markstrahlen gebildet werden, die das sekundäre Xylem und das Phloem verbinden.
Sekundäre Veränderungen treten nicht nur im zentralen Zylinder auf. Aufgrund der Zunahme der Dicke des leitfähigen Gewebes wird die primäre Rinde nach außen gedrückt, zerrissen und zusammen mit dem Endoderm abgestoßen.
Im äußeren Teil des Pericyclus entsteht das Phellogen und lagert das Phellema nach außen und das mehrschichtige Phelloderm nach innen ab. So sind die Wurzeln von Dikotylen, die eine Sekundärstruktur haben, mit Periderm bedeckt.
In mehrjährigen Wurzeln kommt Phellogen tief in der Wurzel vor. Dann bildet die Wurzel wie der Stamm ein Rhytidom.
Ein wichtiger ökologischer Aspekt des sekundären Wurzelwachstums ist die Fähigkeit der Wurzeln verschiedener Pflanzen derselben Art, natürlich zu wachsen. Wo die Wurzeln miteinander in Kontakt kommen, vereinigen sie sich durch sekundäres Wachstum. Beim Zusammenwachsen der Wurzeln wird die Kontinuität des Leitungssystems zwischen den beiden Fusionspartnern hergestellt. Dank der Verschmelzung von Wurzeln sind viele Bäume miteinander verbunden, und wenn einige von ihnen gefällt werden, bleiben die Stümpfe lange am Leben. Die Verschmelzung von Wurzeln trägt zur schnellen Übertragung von Infektionen von einem Baum auf einen anderen bei.
Wurzeldifferenzierung in Wurzelsystemenäußert sich in unterschiedlichen Formen. Von größter Bedeutung ist die Unterscheidung in Haupt-, Seiten- und Adventivwurzeln.
In einer besonderen Gruppe werden die Wurzeln unterschieden, die in den apikalen Meristemen höherer Sporenpflanzen entstehen. Es wird angenommen, dass sie die archaischste Gruppe von Wurzeln sind und die größte Ähnlichkeit mit den Primärwurzeln bewahrt haben, die Teil eines einzigen Verzweigungssystems aus oberirdischen und unterirdischen Achsen waren. Die in dieser Gruppe enthaltenen Wurzeln werden als apicogen bezeichnet.
Basierend auf der Reihenfolge des Auftretens von Wurzelsystemen im Verlauf der Evolution und der Zugehörigkeit zu großen systematischen Gruppen können wir drei skizzieren Evolutionsstufe der Wurzelsysteme.
I. Wurzelsysteme höherer Sporen werden ausschließlich durch apigene Wurzelsysteme repräsentiert, die sich sukzessive gegenseitig ersetzen, was der Hauptmechanismus der Erneuerung in Wurzelsystemen ist. Die Verzweigung erfolgt apikal oder lateral, und es ist möglich, dass beide Verzweigungsarten unabhängig voneinander entstanden sind. Es gibt keine Metamorphosen und Wurzeln mit einer engen Spezialisierung.
P. Das Wurzelsystem von Gymnospermen zeichnet sich durch das Vorhandensein einer Hauptwurzel aus, die mit dem Auftreten eines Samens und eines bipolaren Embryos mit einer embryonalen Wurzel in Verbindung gebracht werden kann. Nicht nur Seitenwurzeln untergeordneter Ordnungen wurden in den Erneuerungsmechanismus einbezogen, sondern auch rhizogene Adventivwurzeln (d. h. Adventivwurzeln auf Wurzeln, Adventivwurzeln auf Sprossen erhielten bei Gymnospermen keine merkliche Entwicklung). Spezialisierte und metamorphosierte Wurzeln sind selten.
Sh. Das Wurzelsystem der Angiospermen ist durch eine starke Entwicklung von Adventivwurzeln sowie Metamorphosen und Wurzeln mit enger Spezialisierung gekennzeichnet. Update-Mechanismen sind vielfältig und effizient.
Metamorphosen von Wurzelsystemen
Die Metamorphose von Wurzelsystemen ist eine Reihe von anatomischen und morphologischen Merkmalen, die phylogenetisch entwickelt und erblich festgelegt und durch eine Änderung der physiologischen Funktionen verursacht werden. Die Metamorphose (Metamorphose, griech. Verwandlung) ist das Ergebnis von Veränderungen, die auf genetischer Ebene stattfinden und zu einer morphologischen und funktionellen Umstrukturierung eines Organs, Organsystems oder Organismus als Ganzes führen.
I. Metamorphosen im Zusammenhang mit der Ausführung atypischer Funktionen:
1) Atmungswurzeln;
2) Luftwurzeln von Epiphyten;
3) gestelzte Wurzeln;
4) Sicherungswurzeln.
P. Metamorphosen im Zusammenhang mit Überfunktion:
1) Hackfrüchte;
2) Wurzelzapfen;
3) Wurzelknollen;
4) Mykorrhiza.
Hackfrüchte sind charakteristisch für zweijährige zweikeimblättrige Pflanzen. Dies ist ein vorläufiger Name. Sie entwickeln sich aus vegetativen Organen und haben nichts mit Früchten zu tun. Der untere Teil des Stiels und die Wurzel nehmen an der Bildung der Wurzelfrucht teil.
Im ersten Lebensjahr vegetiert die Pflanze, es bildet sich eine Hackfrucht. Im zweiten Jahr bilden sich Blüten, Früchte und Samen. Damit ist die Hackfrucht ein Organ der Stoffversorgung, das es der Pflanze ermöglicht, die Ruhephase zu verlegen und die Entwicklung für das nächste Jahr abzuschließen.
Wurzeltypen:
1. Monokambial.
Die Anfangswurzel ist ein Diarch-Axialzylinder. Die Bildung einer Wurzelpflanze beginnt mit sekundären Veränderungen, die mit der Initiierung und dem Betrieb eines Kambialrings und weiterer Parenchymisierung verbunden sind.
Wir wissen, dass das Kambium das sekundäre Xylem nach innen und das sekundäre Phloem nach außen legt. Anatomische Unterschiede beziehen sich auf den Entstehungsort des Speichergewebes. Kommt es im Phloem zu einer starken Entwicklung des Parenchyms, bildet sich ein Phloemtyp der Wurzelfrucht - der Karottentyp. Wenn in Xylem - Xylem-Typ - Rübentyp.
2. Polykambiale Art von Hackfrüchten.
Ein typischer Kambialring wird gebildet, der Rest sind weitere perizyklischen Ursprungs. Sie können bis zu 25. Diese zusätzlichen Kambiumringe bilden offene kollaterale Leitbündel. Je weiter der Ring vom ersten Kambialring entfernt ist, desto mehr Speicherparenchym produziert er. Ein Beispiel für eine polykambiale Wurzelpflanze sind Rüben.
Bei mono- und polykambialen Hackfrüchten stirbt die Primärrinde früh ab und blättert ab. Das Deckgewebe ist das Periderm.
Somit ist ein gemeinsames Merkmal aller Hackfrüchte das Vorhandensein einer großen Menge an Speicherparenchym, das mit leitfähigem Gewebe durchdrungen ist. Die enge Verbindung zwischen Leit- und Speichergewebe wird durch verschiedene Modifikationen des Sekundärwachstums erreicht.
Bei manchen Pflanzen werden Speicherwurzeln aus Seiten- und Adventivwurzeln in Form von Wurzelzapfen gebildet. Wurzelzapfen von Monokotylen (Ljubka) sind kambial, haben eine Primärstruktur; Dicots - Kambium, haben eine Sekundärstruktur. Beispiel: Dahlien-Wurzelzapfen vom Phloem-Typ.
Wurzelzapfen bilden auf der gesamten Oberfläche Adventivwurzeln und im oberen Teil Adventivknospen und sind daher Organe der vegetativen Vermehrung.
Es ist bekannt, dass das Pflanzenwachstum meistens durch eine unzureichende Versorgung mit festem Stickstoff begrenzt wird. Stickstoff in Form des stabilen Moleküls N 2 macht 80 % der Atmosphäre aus. Vor der Fixierung (Reduktion von N 2 zu Ammoniak NH 3) muss dieses Molekül gespalten werden. Die Stickstofffixierung wird von einigen frei lebenden Bakterien durchgeführt, die organische Bodensubstanz verbrauchen. Einige Bakterien siedeln sich an den Wurzeln höherer Pflanzen an. Die Pflanze "lockt" Bakterien mit organischen Wurzelsekreten an. Und Mikroorganismen wiederum versorgen die Pflanze mit festem Stickstoff. Diese für beide Seiten vorteilhafte Verbindung zweier Organismen wird als Symbiose bezeichnet. Am häufigsten bilden die Wurzeln Assoziationen mit stickstofffixierenden Bakterien der Gattung Rhizobium. Mehr als 60 Familien dikotyler und monokotyler Pflanzen sind in der Lage, mit Bakterien eine Symbiose einzugehen. Jede Pflanzenart hat ihr eigenes symbiotisches Rhizobium. Die gegenseitige Erkennung einer Pflanze und eines Bakteriums erfolgt durch Anheften eines speziellen Proteins - eines Lektins, das sich auf der Oberfläche von Trichoblasten befindet - an ein bestimmtes Bakterium. Wurzelhaare unter dem Einfluss von Wachstumshormonen, die von Bakterien ausgeschieden werden, werden deformiert. Bakterien gelangen in diese verdrehten Zellen. In den Zellen vermehren sich die Bakterien und bilden einen von einer Zahnfleischhülle umschlossenen Infektionsfaden. Dieser Faden dringt tief in die Wurzel ein. Das Ergebnis eines solchen Eindringens von Bakterien ist eine starke Proliferation von Wurzelmesodermzellen, die zur Bildung von Knötchen führt. Der äußerste Teil des Knötchens bleibt lange Zeit eine meristematische Zone, und im inneren Teil des Knötchens befinden sich Bakterien. Das Bakteriengewebe selbst wird durch kleine Zellen mit einem viskosen Zytoplasma und einer großen Anzahl von Bakterien dargestellt. Das Bakteriengewebe ist von Gefäßbündeln umgeben, die mit dem zentralen Zylinder der Wurzel verbunden sind. Diese. Es gibt ein Transportsystem für den Austausch von Nährstoffen zwischen Bakterien und Pflanzen. Stickstofffixierende Bakterien wirken sich günstig auf die Stickstoffversorgung von Pflanzen und die Bodenfruchtbarkeit aus.
Die physiologische Aktivität der Wurzel, die mit der Versorgung der Pflanze mit Wasser und Mineralien verbunden ist, kann durch ihre symbiotische Verbindung mit einem bestimmten Pilz verstärkt werden. So eine enge, für beide Seiten vorteilhafte Symbiose von Wurzeln und Pilzen ist Mykorrhiza. Mykorrhiza ist in den meisten Gefäßpflanzen bekannt. Nur wenige Blütenfamilien bilden es nicht oder selten, zum Beispiel Kohl, Segge.
Mykorrhizapilze spielen eine Schlüsselrolle in der mineralischen Ernährung von Gefäßpflanzen. Bedeutende Rolle der Mykorrhiza bei der Absorption und Übertragung von Phosphor.
Während des bilateralen Transports (Bewegen von Phosphaten von Pilzen zu Pflanzen und Kohlenhydraten in die entgegengesetzte Richtung) treten Verlust und Absorption in der apoplastischen Zone zwischen den Plasmamembranen von Pflanze und Pilz auf.
Mykorrhizapilze nutzen P-Quellen, die Pflanzen normalerweise nicht zur Verfügung stehen. Mykorrhiza trägt auch zur Aufnahme von Zn, Mn, Cu durch Pflanzen bei. Diese Elemente im Boden sind schwach mobil, und daher bilden sich um die Wurzelhaare herum schnell entleerte Zonen. Das Hyphennetz der Mykorrhizapilze erstreckt sich mehrere Zentimeter von jeder Wurzel und vergrößert dadurch die Absorptionsfläche der Wurzeln.
Es gibt folgende Arten von Mykorrhiza:
1) Ektomykorrhiza: Der Pilz bedeckt die gesamte Wurzelspitze mit einer dicken Hülle und dringt in die Interzellularräume ein;
2) Endomykorrhiza: Der Pilz dringt in die Zellen ein.
Ektomykorrhiza ist charakteristisch für bestimmte Baum- und Strauchfamilien (Kiefer, Weide, Buche). Ektomykorrhiza wird an jungen, noch nicht suberisierten Wurzeln gebildet. Ektomykorrhiza-Wurzeln sind kurz, verzweigt und sehen geschwollen aus.
In solchen Wurzeln wird die Entwicklung von Wurzelhaaren unterdrückt und das Volumen des apikalen Meristems und der Wurzelkappe kann reduziert werden. Die Funktion der Wurzelhaare wird von Hyphen übernommen.
Endomykorrhiza-Wurzeln haben eine ähnliche Form wie gewöhnliche Wurzeln, unterscheiden sich jedoch von ihnen durch eine dunklere Farbe. Pilzhyphen dringen in die Zellen der Wurzelrinde ein, wo sie Bläschen (Schwellungen) und Arbuskeln (baumartige Äste) bilden. Daher wird Endomykorrhiza oft als vesikulär-arbuskulär bezeichnet. Unter bestimmten Bedingungen können Pilzhyphen von Zellen „verdaut“ werden. Wurzelhaare an den Wurzeln bleiben erhalten. Dies ist die häufigste Art von Mykorrhiza. Es kommt in etwa 80 % aller Gefäßpflanzen vor, nämlich in krautigen Pflanzen, einschließlich Getreide; an Sträuchern, Bäumen (Apfelbäume, Ahorne ...). In einer jungen Pappelpflanze wird beispielsweise eine vesikulär-arbuskuläre Endomykorrhiza gebildet, und mit zunehmendem Alter entwickelt sich eine Scheiden-Ektomykorrhiza.
Die Heidekrautgewächse Ericaceae sind durch erikoid-arbutoide Mykorrhiza gekennzeichnet. Der Pilz bildet eine starke Ansammlung von Hyphen um die Wurzel, und dünne Seitenhyphen dringen in die Zellen der Wurzelrinde ein. Diese Mykorrhiza-Pilze reduzieren die Bodentoxizität für Heiden erheblich und ermöglichen es diesen Pflanzen, sich in sauren, nährstoffarmen Böden anzusiedeln.
Es gibt noch eine andere Art - Orchideen-Endomykorrhiza. Orchideensamen keimen in der Natur nur in Gegenwart geeigneter Pilze.
Im Allgemeinen sind Mykorrhiza-Assoziationen unter natürlichen Bedingungen eher die Regel als die Ausnahme.
Eine Untersuchung der Überreste der ältesten Gefäßpflanzen zeigte, dass Endomykorrhiza bei ihnen nicht weniger häufig vorkam als bei modernen Pflanzen. Dies ließ vermuten, dass die Entstehung von Mykorrhiza im Verlauf der Evolution einer der Faktoren sein könnte, die es Pflanzen ermöglichten, das Land zu besiedeln. Unter Berücksichtigung der schwachen Bodenentwicklung während dieser Besiedlung kann davon ausgegangen werden, dass Mykorrhizapilze die Grundlage der mineralischen Ernährung der Pflanzen waren. Moderne Pionierpflanzen mit Mykorrhiza haben bessere Überlebenschancen auf mageren Böden.
Pflanzen mit mykotropher Ernährung sind widerstandsfähiger gegen das Vorhandensein von Schwermetallen in der technogenen Umgebung.
Ein paar Worte zu den Wurzeln tropischer Regenwaldpflanzen. Häufiger als gestelzte, brettartige Stützwurzeln. Die Wurzeln ähneln Brettern, die mit ihren oberen Enden gegen einen Baum gelehnt sind. Während Stelzenwurzeln Adventivwurzeln sind, die am Stamm entstehen und nach unten wachsen, ist die Plankenwurzelbildung mit Seitenwurzeln verbunden, die direkt unter der Bodenoberfläche wachsen. Seitenwurzeln entwickeln vertikale Auswüchse mit dreieckigem Querschnitt, die an den Stamm des Baumes angrenzen.
Das Holz der Plankenwurzeln ist oft härter als das übrige Holz desselben Baumes, und oft sind sie mit einer dünnen Rindenschicht bedeckt. Den im Querschnitt sichtbaren Wachstumsringen dieser Wurzeln nach zu urteilen, handelt es sich zunächst um normale, rundliche Wurzeln, die dann aber an der Oberseite eine sehr starke Verdickung entwickeln. Die Anzahl der Plankenwurzeln an einem Baum variiert zwischen 1 und 10, aber in den meisten Fällen sind es drei oder mehr. In den Tropen macht sich die Bildung von Plankenwurzeln besonders bei Bäumen bemerkbar, die in Sumpfwäldern und schlecht entwässerten Böden wachsen. Oft werden brettförmige Wurzeln für industrielle Zwecke verwendet. So werden die plankenartigen Wurzeln des in Indonesien wachsenden Baums Koompassia excelsa zur Herstellung von Esstischen verwendet.
DIE FLUCHT
Der Spross ist wie die Wurzel das wichtigste vegetative Organ der Pflanze. Der Spross erfüllt die Funktion der Lufternährung, kann eine Reihe anderer Funktionen erfüllen und ist zur Metamorphose fähig.
Ein vegetativer unveränderter Spross besteht aus einem axialen Teil - einem Stamm; Blätter und Knospen. Das wichtigste äußere Merkmal, das den Spross von der Wurzel unterscheidet, ist die Belaubung des Sprosses.
Der Abschnitt des Stängels, der mit der Entstehung von Seitenorganen (Zweige, Blätter usw.) verbunden ist - Knoten. Der Abschnitt des Stammes zwischen benachbarten Knoten Internodien. Innerer Winkel zwischen Blatt und Stängel Blattachsel. Jedes sich wiederholende Element des Sprosses (ein Knoten mit einem Blatt und einer Knospe in der Blattachsel und einem Internodium) bildet sich metamer. Der Spross besteht somit aus einer Reihe von Metameren, d.h. hat eine metamere Struktur.
In den frühen 90er Jahren des 20. Jahrhunderts schlugen amerikanische Wissenschaftler anstelle der Konzepte eines Knotens und eines Internodiums das Konzept von - Stammelement. Das Stängelelement ist der Abschnitt des Stängels zwischen den Rändern der Blattanlagen. Offenbart an der Triebspitze, wo noch kein Internodium vorhanden ist. Diese. der neue Begriff „Stammelement“ entspricht dem Begriff „Metamer“.
Die Struktur des Sprossapikalmeristems ist komplexer als die der Wurzel. Tatsächlich bildet das apikale Meristem im Spross nicht nur Zellen von Primärgeweben, sondern ist auch an der Bildung von Blattanlagen und Achselknospen beteiligt, die sich dann zu Seitensprossen entwickeln. Dieses Meristem ist nicht durch eine Struktur wie eine Wurzelkappe geschützt.
Die Arten von Spitzen sind die gleichen wie in der Wurzel: monoinitial, zonal, gestuft. Die Anzahl der Ebenen ist größer als an der Wurzel.
Evolutionär primitive monoinitiale Spitze. Ein Beispiel für diese Art von Apex findet man in Schachtelhalmen. Eine einzelne große längliche apikale Zelle ist stark vakuolisiert. Es teilt sich in Richtungen parallel zu seinen Oberflächen (mit Ausnahme der äußersten Oberfläche). Die nächsten abgeleiteten Zellen sind ebenfalls stark vakuolisiert, aber wenn sie sich teilen, erscheinen kleine Zellen mit dichten Protoplasten. Solche Zellen befinden sich entlang der Peripherie der Stammspitze, wo Blattanlagen abgelegt werden.
Die vegetative Spitze des Triebs der meisten Blütenpflanzen ist wie eine Tunika aufgebaut - ein Körper - zwei Zonen, die sich in den Ebenen der Zellteilung unterscheiden. Tunika(lat. - Schale) umfasst die äußeren Zellschichten, die sich antiklinal (senkrecht zur Oberfläche) teilen und für eine Vergrößerung der Oberfläche sorgen. Rahmen(lat. - Körper) - die zentrale Gruppe von Zellen unter der Tunika teilt sich in verschiedene Ebenen und bildet das Hauptvolumen des Triebs. Der Körper und jede Schicht der Tunika hat ihre eigenen Initialen. In der Tunika sind die Initialen in der Mitte entlang der Achse platziert. Zweikeimblättrige haben hauptsächlich eine zweilagige Tunika, Einkeimblättrige haben eine einlagige, seltener eine zweilagige Tunika.
Während das Konzept einer Tunika-Hülle zu unserem Verständnis der Lage und des Wachstums von Zellen im apikalen Meristem selbst beiträgt, hilft die Erkennung verschiedener Zellzonen in angrenzenden Bereichen, den Beitrag des Meristems zur Differenzierung von Geweben und Organen aufzudecken der Schuss. Das Vorhandensein von Zonen mit unterschiedlichen zytologischen Eigenschaften im apikalen Meristem ist Zytohistologische Zonierung.
Der Korpus entspricht einer Region von Zellen, die als Zone der zentralen Mutterzellen. Die Zellen dieser Zone sind stark vakuolisiert, ein Merkmal, das mit einer relativ geringen Rate an mitotischer Aktivität verbunden ist. Die Zone der zentralen Mutterzellen ist von einem peripheren Meristem umgeben, das teils aus der Tunika, teils aus dem Corpus entspringt. Das Protoderm-Histogen stammt aus der äußeren Schicht der Tunika. Das periphere Meristem stammt von den lateralen Ableitungen der apikalen Initialen und von den zentralen Mutterzellen. Das Kernmeristem entsteht durch Teilungen entlang der Peripherie der zentralen Mutterzellen in einer sogenannten Schicht Übergangszone. Die Zellen der peripheren Zone haben dichte Protoplasten, daher haben sie eine hohe mitotische Aktivität.
An bestimmten Stellen mit besonders hoher Aktivität werden Blattknollen (oder Primärknollen) abgelegt.
Die Entstehung einer neuen Blattanlage ist mit einer Richtungsänderung der Zellteilung verbunden, der eine Änderung der Ausrichtung der Mikrotubuli in den peripheren Schichten des Zytoplasmas vorausgeht. Auch der osmotische Druck beeinflusst das Wachstum der Primordien.
Während der Phase des aktiven Wachstums bildet das Sprossspitzenmeristem so schnell Blattanlagen aus, dass sich die Knoten und Internodien zunächst nicht unterscheiden. Allmählich beginnen die Bereiche zwischen den Blattschichten zu wachsen, sich zu dehnen und die Form von Internodien anzunehmen. Gleichzeitig werden die Blattanhaftungsbereiche zu Knoten geformt.
Unten, wo die Rudimente der Blätter erscheinen, wird eine Vakuolisierung beobachtet. Dies zeigt den Beginn der Differenzierung der primären Rinde und des Marks an. Eine starke Vakuolisierung in der primären Rinde und im Mark weist auf das Vorhandensein einer meristematischen Zone hin, die als Restmeristem bezeichnet wird. Es sieht aus, als wäre es ein Rest des apikalen Meristems, der zwischen differenzierteren Geweben erhalten geblieben ist. Unterhalb der Ablagestelle des Blattrudiments im Restmeristem werden durch Längsteilungen ohne anschließendes Wachstum der Zellen in die Breite etwas verlängerte Zellen des ersten Procambiums gebildet.
In den nächsten Entwicklungsstadien differenzieren sich immer mehr Prokambialstränge im Restmeristem aus. Nachdem sich alle Leitbündel einer bestimmten Stammebene gebildet haben, differenziert sich das Restmeristem zum interfaszikulären Parenchym. An den Knoten wird ein Teil des Restmeristems zum Blattschlitzparenchym.
Das Prokambium kann in einem durchgehenden Ring oder in getrennten Strängen verlegt werden. Wenn das Procambium in einem durchgehenden Ring liegt, a trägerlose (nicht gebündelte) Art der Stammstruktur. Wenn das Procambium in getrennten Strängen verlegt wird, a Balkentyp der Stammstruktur. Daher befinden sich primäre leitfähige Gewebe im Stamm auf unterschiedliche Weise: im ersten Fall in einem durchgehenden Ring, im zweiten Fall in separaten leitfähigen Bündeln, die durch Abschnitte des Hauptparenchyms - Kernstrahlen - getrennt sind.
In den kollateralen Leitbündeln erscheint das primäre Phloem im äußeren Teil des Procambialstrangs und das primäre Xylem im inneren Teil. Die anschließende Differenzierung des Phloems erfolgt zentripetal: Neue Elemente des Phloems erscheinen näher an der Mitte des Stammes. Xylem differenziert in die entgegengesetzte Richtung - zentrifugal.
Die gesamte Stele oder der zentrale Teil des Stammes besteht aus leitfähigem Gewebe, dem Kern (manchmal kollabiert er) und dem Perizykel. Außerhalb des Perizykels befindet sich der primäre Cortex. Die Zusammensetzung des primären Kortex (und er wird aus dem peripheren Meristem gebildet) umfasst Chlorenchym, manchmal sekretorische Elemente. Die äußerste Schicht des peripheren Meristems bildet kein Exoderm wie in der Wurzel, sondern differenziert sich zu Collenchym, das entweder in Form von Strängen oder in einem durchgehenden Ring (je nach Art der Stängelstruktur: gebündelt oder bündellos) vorliegt. Die innerste Schicht der primären Rinde ist das Endoderm. Aber im Stamm hat es keine regulatorische Funktion wie in der Wurzel. Stärkekörner lagern sich im Endoderm ab und werden daher als stärkehaltige Hülle bezeichnet. Manchmal enthält es Calciumoxalatkristalle.
Das Pericyclus verliert schnell seine meristematische Aktivität und wandelt sich in Sklerenchym um, das je nach Art der Stammstruktur in einer durchgehenden Schicht oder in Strängen auftritt.
Den Kern bilden dünnwandige Parenchymzellen, in denen Reservenährstoffe abgelagert sind. Es gibt Idioblastenzellen, die mit Kristallen, Schleim usw. gefüllt sind. Der periphere Teil des Kerns wird genannt perimedulläre Zone(Griechisch peri - herum, Medulla - Kern). Die Zellen der Perimedullärzone sind kleiner und behalten ihre vitale Aktivität lange bei.
Bei Monokotylen ist der Anfangstyp der Bündeltyp der Stammstruktur, d.h. Procambium liegt in Strängen. Aber das sekundäre Meristem – das Kambium – entsteht nie, und daher gibt es keine sekundären leitfähigen Gewebe. Selbst bei Verdickung wird der Stamm nur von Primärgeweben gebildet. Der dicke Stamm von Monokotylen (Palmen, Aloe) entwickelt sich aufgrund der Teilung und Streckung der Zellen des Hauptparenchyms. Dieses Wachstum wird als diffuses sekundäres Wachstum bezeichnet.
Bei Monokotylen sind der primäre Kortex und der zentrale Zylinder nicht wie bei Dikotylen durch eine klare Grenze getrennt, und die Stele erhebt sich unter der Epidermis.
Monocots zeichnen sich durch eine diffuse Art der Stammstruktur aus, wenn geschlossene Leitbündel über den gesamten Stamm verstreut sind. Bei der Dehnung der Internodien werden die Primärelemente des Phloems und Xylems (Protophloem und Protoxylem) gedehnt und zerstört, wodurch auf der Xylemseite des Bündels ein großer lufttragender Hohlraum entsteht. Daher wurde in Laborstudien, als wir uns die Arten von Bündeln an einer permanenten Präparation eines Querschnitts eines Maisstängels ansahen, klar, dass ein reifes Leitbündel zwei breite Metaxylemgefäße, eine Lufthöhle und ein Metaphloem umfasste. Jedes Bündel ist von einer Auskleidung aus Sklerenchymzellen umgeben.
Bei Monokotyledonen stirbt oft der Stängelkern ab und an seiner Stelle bildet sich ein Hohlraum. Ein solcher hohler Stamm mit gut definierten Knoten wird genannt Stroh. Solomina zeichnet sich durch eine starke Entwicklung von mechanischem Gewebe aus, das an der Peripherie des Stiels einen Ring bildet. In reifen Pflanzen wird Chlorenchym zerstört und das Stroh erhält eine für Sklerenchym charakteristische goldgelbe Farbe. Im Allgemeinen sind Monokotyledonen durch eine starke Sklerifikation des Stammes gekennzeichnet, wenn bis zu 30% aller Gewebe Sklerenchym sind, und es kann zu einer teilweisen Verholzung der Membranen lebender Parenchymzellen kommen.
Im Stamm von Dikotylen ist das Sekundärwachstum das Ergebnis der Aktivität des Kambiums. Kambium erscheint am Ende des ersten Jahres des Sprosswachstums und trägt zur Verdickung des Stängels bei. Das Kambium entspringt teilweise aus dem Procambium in den Leitbündeln und teilweise aus dem Interfaszikularparenchym. Je nach Entstehungsort werden Abschnitte des Kambiums als faszikuläres und interfaszikuläres Kambium bezeichnet. Zukünftige Kambialzellen in den interfaszikulären Zonen werden lange vor Beginn der Kambiumaktivität bestimmt. Das Kambium wird zuerst in den an den Fasciculus angrenzenden Zellen gebildet und breitet sich dann tief in den Markstrahl aus. Dadurch bilden das interfaszikuläre und das faszikuläre Kambium einen durchgehenden Kambiumring im Stamm.
Es gibt drei Arten von sekundärem Wachstum:
1) Aristolochia chircason-Typ.
Der Anfangsbuchstabe ist der Balkentyp der Stammstruktur. Primäre Leitbündel sind durch breite Kernstrahlen getrennt. Das gebündelte Kambium bildet sekundäres leitfähiges Gewebe, und das interfaszikuläre Kambium bildet das Strahlungsparenchym. Diese Balkenstruktur bleibt erhalten.
2) Sonnenblumentyp Helianthus.
Der Balkentyp der Struktur ist auch der ursprüngliche. Sekundäre leitfähige Gewebe werden jedoch sowohl vom faszikulären als auch vom interfaszikulären Kambium gebildet. Die vom interfaszikulären Kambium gebildeten sekundären Gefäßbündel unterscheiden sich von den primären durch kleinere Größen und das Fehlen von mechanischem Gewebe perizyklischen Ursprungs über dem Bündel. Bei längerer Einwirkung des Kambiums entstehen immer mehr Nebenbündel. Sie wachsen, verschmelzen miteinander und bilden einen kontinuierlichen Ring aus sekundären leitfähigen Geweben.
3) Tilia-Linden-Typ.
Der anfängliche Typ ist der balkenlose Typ einer Stammstruktur. Der gemeinsame Kambialring bildet sekundäres leitfähiges Gewebe. Aber von Zeit zu Zeit bildet das Kambium Parenchymzellen, die sich weiter zu Sekundärstrahlen differenzieren.
Stammstruktur Stauden
Bei holzigen und strauchigen Dikotyledonen sowie bei Nadelbäumen dauert die sekundäre Verdickung viele Jahre an, und bei einigen Arten erreichen die Stämme einen Durchmesser von mehreren Metern. Verdickungen sind mit der Aktivität eines langfristig funktionierenden Kambiums und teilweise Phellogens verbunden - zwei sekundäre seitliche Meristeme. Topographisch lassen sich im Stängel eines mehrjährigen Gehölzes drei Hauptteile unterscheiden: Rinde, Holz und Mark. Die Grenze zwischen Rinde und Holz verläuft entlang des Kambiums.
Kambiumzellen sind im Gegensatz zu typischen meristematischen Zellen immer stark vakuolisiert. Der Zustand des Zytoplasmas von Kambialzellen ändert sich mit den Jahreszeiten.
In der Kambialzone oder dem Kambialring finden sich zwei Formen von Kambialzellen: in vertikaler Richtung verlängerte spindelförmige Zellen (Initialen) und in horizontaler Richtung verlängerte kugelförmige Zellen - Strahlinitialen. Wenn die Spindelzellen in regelmäßigen horizontalen Reihen angeordnet sind, wird das Kambium als Stufenkambium bezeichnet. Wenn bei der Anordnung spindelförmiger Zellen keine solche Reihenfolge vorhanden ist, ist das Kambium nicht gestuft.
Langleinenkambium führt zu Langleinenholz und Langleinen-Sekundärphloem. Nicht-gestuftes Kambium führt zu nicht-gestuftem Holz und nicht-gestuftem sekundärem Phloem. Langleinenhölzer und Phloeme gelten evolutionär als höher spezialisiert als nicht geschichtete Hölzer.
Mit der Verdickung des Stiels nimmt auch der Umfang der Schichten des Kambiums zu. Möglich wird dies durch die Teilung der Kambialzellen in radialer Richtung.
Unter den Kambialzellen ist die mittlere Anfangsschicht isoliert. Als Ergebnis der Teilung bilden seine Zellen Derivate im Inneren und dann im Inneren Richtung nach außen. Für den gleichen Zeitraum werden 2-3 Zellen nach innen und nur eine Zelle nach außen abgelagert. Dadurch entsteht mehr Holz als Bast.
Balkenzellen (Initialen) bilden Kernparenchymstrahlen: primäre, die sich vom Kern bis zur Rinde erstrecken; und sekundäre Kernstrahlen sind kürzer, weil entstehen aus später entstandenen Holzstrahlzellen.
Kernträger können einreihig und mehrreihig sein; homogen (d. h. aus denselben Zellen bestehend) und heterogen (der Strahl enthält Zellen, die sich in der Struktur unterscheiden).
Bedeutung der Kernstrahlen : Es ist ein Speicherort für Nährstoffe (Stärke, Lipide), die während der Ruhezeit (z. B. im Winter) eine Rolle spielen.
Die Kernstrahlen sorgen für einen radialen Transfer von Wasser und Mineralsalzen vom Xylem entlang des Apoplasten zum Kambium und sekundären Phloem und in der entgegengesetzten Richtung entlang des Nährsymplasten.
Und schließlich findet der Gasaustausch über die Interzellularräume statt.
Holz
Holz wird durch das Kambium gebildet und wächst in zentrifugaler Richtung. Da In unserer Zone arbeitet das Kambium periodisch: Es beginnt im Frühjahr zu arbeiten und hört am Ende des Sommers auf, dann ist eine konzentrische Schichtung im Holz sichtbar. Im Frühjahr, zu Beginn der Vegetationsperiode, bildet sich Frühholz. Seine Gefäße sind breit, dünnwandig und können eine beträchtliche Menge Wasser transportieren. Wasser wird benötigt, um das Wachstum zu initiieren, insbesondere um die Größe neuer Zellen zu erhöhen (z. B. sich entwickelnde Blattzellen). Spätholz enthält weniger Gefäße. Sie sind schmaler und dicker. Der Übergang vom Früh- zum Spätholz des gleichen Jahres kann fließend sein. Allerdings ist die Grenze von Spätholz zu Frühholz einjährig nächstes Jahr immer scharf und im Querschnitt gut sichtbar. Dadurch entsteht ein einjähriger Bewuchs (Jahresring oder Jahresschicht).
Nach der Platzierung der Gefäße im jährlichen (jährlichen) Wachstum werden folgende Holzarten unterschieden.
Wenn sich die Gefäße hauptsächlich im Frühholz befinden und im Spätholz fehlen oder nur kleine Gefäße platziert sind, wie Eiche, Esche, ist dies der Fall ringförmiges Holz.
Wenn die Gefäße gleichmäßig über den Jahreszuwachs verteilt sind, nimmt ihr Durchmesser zwar im Spätholz ab, dies ist jedoch der Fall verstreutes Gefäßholz. Gefunden in Birke, Ahorn, Pappel.
Auch zwischen diesen Holzarten gibt es Übergangsformen.
Da Holz leitende, mechanische und Speicherfunktionen erfüllt, umfasst es neben Xylemelementen auch mechanisches Gewebe in Form von Sklerenchym und Speichergewebe in Form des Hauptparenchyms. Die Struktur aller anatomischen Elemente, ihre Anzahl und Anordnung verleihen dem Holz verschiedener Pflanzen spezifische Merkmale, die für eine bestimmte taxonomische Gruppe charakteristisch sind.
Evolutionär weiter fortgeschrittene Pflanzen haben zum Beispiel ein spezialisiertes mechanisches Gewebe (Sklerchym). Evolutionär primitive zweikeimblättrige (Magnolie) haben keine Holzfasern. Die mechanische Rolle spielen in diesem Fall Tracheiden.
Im Holz einiger Pflanzen gibt es Übergangsformen von Tracheiden zu Libriform (Libriform sind spezialisierte mechanische Elemente des Holzes, die aus Prosenchymzellen bestehen, die an den Enden mit dicken verholzten Schalen zugespitzt sind). Es gibt auch eine Cloisonne libriform. Seine ursprüngliche spindelförmige Zelle mit dicken vertikalen Membranen ist quer in einzelne kurzlebige Zellen unterteilt. Solche histologischen Elemente sind morphologisch und funktionell holzigen Parenchym nahe.
Holzparenchym spielt eine Speicher- und teilweise leitende Rolle. Parenchymzellen leben. Das Holzparenchym ist in horizontalen und vertikalen Reihen angeordnet. Die horizontalen Reihen bilden die Kernstrahlen. Wir haben ihre Eigenschaften bereits berücksichtigt. Die vertikalen Reihen bilden das holzige Parenchym selbst.
Wenn das Holzparenchym während des Jahreswachstums verstreut ist, wird dies diffuses Parenchym. Typisch für Linde, Eiche, Birnbaum.
Wenn sich das Holzparenchym um die Gefäße befindet, wird dies paratracheales Parenchym. Esche, Ahornholz.
An der äußeren Grenze befindet sich der Jahreszuwachs terminales Parenchym. Beispiel: Weiden-, Lärchen-, Magnolienholz.
Die Besonderheit seiner Struktur ist das Fehlen von Blutgefäßen. Die Trachealelemente sind nicht perforiert und werden durch Tracheiden dargestellt, die sowohl leitende als auch mechanische Rollen übernehmen.
Koniferen-Tracheiden zeichnen sich durch große umrandete Poren mit einem Torus aus. Tracheiden sind lang mit spitzen Enden. Frühe Tracheiden sind groß und dünnwandig. Spätere Tracheiden sind radial abgeflacht, dickwandig.
Wir sehen, dass das Holz von Gymnospermen und Dikotylen signifikante strukturelle Unterschiede aufweist. Daher wird es in zwei Hauptgruppen unterteilt. Das Holz von Gymnospermen wird als weich und das Holz von Dikotylen als hartes Holz klassifiziert. Diese Begriffe "hart" und "weich" sind nicht notwendigerweise auf den Dichte- und Härtegrad beschränkt. Die Unterschiede hängen, wie wir bereits gesagt haben, mit strukturellen Merkmalen zusammen.
In geneigten oder gebogenen Stämmen entsteht reaktives Holz. Seine Bildung wird durch die Tendenz von Ästen und Stämmen verursacht, den Belastungen zu widerstehen, die auftreten, wenn sie geneigt sind, d.h. seine Bildung ist mit dem Begradigungsprozess dieser Pflanzenteile verbunden.
Die wichtigsten Einflussfaktoren auf die Entwicklung von reaktivem Holz sind Gravitationskräfte und die Verteilung von körpereigenen Wachstumsstimulanzien.
Reaktivholz unterscheidet sich sowohl anatomisch als auch chemisch von herkömmlichem Holz. Reaktive Holzzellen werden stärker verholzt und wandverdickt. Bei Nadelbäumen ist es dichter und dunkler als das umgebende Gewebe. Seine Tracheiden sind kürzer als die von normalem Holz.
Die Radioaktivität von Stammholz nimmt von den äußeren Schichten zum Kern hin ab. Es gibt eine gewisse Zunahme der Holzverunreinigung von der Basis des Stammes bis zur Spitze.
Ein ausreichend hoher Gehalt an radioaktiven Stoffen in Rinde und Sprossen weist auf deren äußere Luftkontamination hin.
Bellen
Dieser Begriff umfasst alle Gewebe außerhalb des Kambiums. Dies ist bereits ein sekundäres Bellen, weil. wird von einem sekundären Meristem gebildet - Kambium (im Gegensatz zum primären, das während der Differenzierung von Zellen des primären Meristems gebildet wird).
Die Zusammensetzung des sekundären Kortex umfasst das sekundäre Phloem, Sklerenchym (Bastfasern und Steinzellen) und das Hauptparenchym. Die Menge der mechanischen Elemente wird aufgerufen harter Bast, leitfähige Elemente und das Hauptparenchym - weicher Bast.
In der sekundären Rinde befinden sich sekretorische Zellen und Harzkanäle. Die Hauptfunktion des sekundären Kortex ist die Leitung von Kunststoffsubstanzen sowie die Schutzfunktion.
Das sekundäre Phloem macht einen viel geringeren Anteil des Stammvolumens aus als das sekundäre Xylem. Dies liegt daran, dass das Kambium erstens weniger Phloem bildet als Xylem und zweitens das alte, nicht mehr funktionierende Phloem nach und nach zerkleinert wird.
Aufgrund der Umfangsvergrößerung des Achsenorgans dehnen sich die Parenchymstrahlen aus und ähneln einem Dreieck, dessen Spitze auf das Kambium gerichtet ist. Ein solches Parenchym wird als erweitert bezeichnet (lat. dilatatio - Erweiterung). Diese Anordnung des Parenchyms verhindert, dass der Bast mit zunehmender Dicke des Stiels einreißt.
Bei Nadelbäumen hat das sekundäre Phloem eine einfachere Struktur. Im Bast von Gymnospermen sind histologische Elemente gleichmäßig verteilt und natürlich sind Harzkanäle vorhanden.
Alle Gewebe außerhalb des Phellogens (d. h. das innerste Korkkambium) nehmen kein Wasser und keine Mineralien mehr auf, wenn die Korkzellen suberinisiert werden. Alle diese Periderme mit den Überresten der Rinde (abgestorbene Zellen des Parenchyms und des Phloems) bilden die äußere (äußere) Rinde. Der lebende Teil der Rinde zwischen dem Kambium und der innersten Schicht des Phellogens wird als innere Rinde bezeichnet.
So haben wir, aufbauend auf unseren Erkenntnissen aus dem Thema „Gewebe“, den anatomischen Aufbau des Stammes als Achsenorgan des Sprosses untersucht.
Betrachten Sie nun die Struktur des Seitenorgans des Triebs - des Blattes.
BLECH
Das Blatt ist das wichtigste Photosyntheseorgan höherer Pflanzen. Der Aufbau des Blattes und seine Funktionen sind eng miteinander verbunden.
Aus der Gesamtgleichung der Photosynthesereaktionen:
CO 2 + H 2 O chl-l → hv (CH 2 O) n + O 2
wir können daraus schließen, dass 1) Blätter eine CO 2 - und Wasserquelle benötigen; 2) Blätter müssen an die Absorption angepasst sein Solarenergie sie müssen Chlorophyll haben; 3) O 2 wird als eines der Reaktionsprodukte freigesetzt; 4) Kohlenhydrate müssen entweder auf Vorrat gelagert oder zu anderen Teilen der Pflanze transportiert werden. Die Liste ist ein sehr spezialisiertes Gremium, das all diese Anforderungen erfüllt.
Die gesamte Evolution des Blattes als vegetatives Organ folgte dem Entwicklungspfad der Anpassung an die optimale Nutzung des Lichts.
Gemäß der mikrophilen Evolutionslinie gilt das Blatt als Auswuchs des Teloms archaischer Pflanzen vom Rhinophytentyp und behält die strukturelle Struktur des Achsenorgans bei.
Gemäß der makrophilen Evolutionslinie entstand das Blatt durch Abflachung, Platzierung von Telomen in einer Ebene und deren anschließende Verschmelzung zu einem Ganzen. Gleichzeitig ging die Fähigkeit zum langfristigen apikalen Wachstum und zur Verzweigung verloren. Was sind die morphologischen und anatomischen Merkmale des Blattes, das als Ergebnis einer langen Evolution entstanden ist?
Ein erwachsenes Blatt besteht normalerweise aus einer Blattspreite und einem Blattstiel. Blattstiel- ein schmaler stielartiger Teil des Blattes zwischen der Blattspreite und dem Sprossknoten, mit dessen Hilfe das Blatt im Raum ausgerichtet und in Bezug auf das Licht am günstigsten platziert wird. Durch den Blattstiel sind die Blattspreite und der Stängel verbunden. Leitfähiges und mechanisches Gewebe sind im Blattstiel gut entwickelt. Beim Laubfall bildet sich im Blattstiel eine Trennschicht.
Der unterste Teil des Blattes, der mit dem Stiel artikuliert ist, wird genannt Blattbasis. Die Basis des Blattes kann eine Vielzahl von Formen annehmen. Meistens sieht es aus wie eine kleine Verdickung und heißt Blattauflage. Häufiger an der Basis des Blattes gibt es verschiedene Formen und Größe der paarigen seitlichen Auswüchse - Nebenblätter. Nebenblätter können blattförmig sein und als zusätzliche Assimilationsfläche dienen. Nebenblätter können nadelförmig sein und als Pflanzenabwehr wirken.
Es gibt schuppige, häutige Nebenblätter. Evolutionär gesehen kommt es zu einer Reduktion der Nebenblätter, bei evolutionär jungen Pflanzen fallen sie früh ab oder fehlen ganz.
Bei einigen Getreidearten, Sellerie (Umbellat), wächst die Basis des Blattes und bildet eine geschlossene oder offene Röhre - die Blattscheide. Die Blattscheide trägt zur langfristigen Erhaltung des interkalaren Meristems bei und dient als zusätzliche Stütze für den Spross.
Der Hauptteil des Blattes ist die Blattspreite. Aufgrund der ungleichmäßigen Beleuchtung der Blattober- und -unterseite hat die Platte eine dorsoventrale Struktur, d.h. Die Struktur der Oberseite des Blattes, ventral, unterscheidet sich von der Struktur der Unterseite, dorsal. Die Oberseite des Blattes wird ventral genannt, weil. bei der Niere liegt diese Seite innen und ist der Sprossachse zugewandt. Die untere, dorsale Seite des Blattes in der Knospe ist von der Pflanze nach außen gedreht. Somit ist ein Blatt mit dorsoventraler Symmetrie bilateral oder bilateral oder bifazial. charakteristisch für Zweikeimblättrige.
Ein Blatt mit radialer Symmetrie ist äquifazial. Solche verdickten, zylindrischen Blätter findet man bei Crassulaceae. Einseitige oder isolierte einseitige Blätter sind charakteristisch für Getreide, Zwiebeln.
In Betracht ziehen anatomischer Aufbau der Blattspreite.
Wie die Wurzel und der Stängel besteht das Blatt aus Haut-, Leit- und Hauptgewebe. Da Das Blatt hat normalerweise kein sekundäres Wachstum (mit Ausnahme eines leichten Wachstums in den Blattstielen und großen Adern), es behält die Epidermis als Integumentgewebe bei.
Stomata befinden sich meist auf der Blattunterseite. In zweikeimblättrigen Blättern sind Stomata in nicht sichtbarer Reihenfolge über die gesamte Oberfläche des Blattes verstreut. Bei einkeimblättrigen und Nadelblättern sind sie in Reihen parallel zur Längsachse des Blattes verteilt. In der Epidermis einiger Getreidearten befinden sich motorische oder motorische Zellen. Sie sind größer als normale Epidermiszellen. Die Vakuole nimmt fast die gesamte Zelle ein. Mit dem Turgorverlust ziehen sich diese Zellen zusammen und tragen zur Faltung oder Kräuselung des Blattes bei.
Der größte Teil des Hauptgewebes der Blattspreite befindet sich im Mesophyll. Das Mesophyll ist differenziert. Palisadenmesophyll befindet sich auf der Oberseite der Platte, schwammig - nach unten. Blätter mit einer solchen Struktur sind bifazial oder dorsoventral.
Wenn sich das Palisadenmesophyll auf beiden Seiten des Blattes befindet, ist das Blatt einseitig oder isoliert.
Im Mesophyll von Getreide gibt es keine eindeutige Unterscheidung in Palisaden- und Schwammchlorenchym. Mesophyllzellen sind radial um die Bündel angeordnet.
Leitfähige Blattbündel werden als Venen bezeichnet, und das Verzweigungssystem der Venen wird als Venen bezeichnet Venation. Merkmale der Anordnung von Leitbündeln in Blattspreiten bestimmen verschiedene Arten von Adern, die dichotom, parallel, bogenförmig, handförmig und gefiedert sein können.
Je nachdem, wie die Venen höherer Verzweigungsordnungen miteinander verbunden sind, unterscheidet man offene (dichotome) und geschlossene Venen. Bei der dichotomen Aderung verzweigen sich die Adern dichotom, gehen in einem spitzen Winkel ab und erreichen den Rand der Blattspreite, wo sie nirgends in benachbarte Adern übergehen. Bei geschlossener Venation bilden kleine Venen infolge von Anastomosen (Kreuzungen) ein Netzwerk, das die gesamte Platte oder den größten Teil davon durchdringt.
Blätter mit handförmiger und gefiederter Aderung, und diese sind meist zweikeimblättrig, haben eine mittlere, größte Aderung. Die Vene besteht aus primärem Xylem und primärem Phloem, die aus dem Procambium hervorgehen und zu Kollateralbündeln zusammengefasst sind. Zwischen Xylem und Phloem bildet sich oft ein Kambium, das aber nicht funktioniert. Daher sind Xylem und Phloem primär.
Die Bündel bilden im Blatt ein durchgehendes System, das mit dem Leitungssystem des Stammes verbunden ist. Daher ist das Xylem immer zur morphologischen Oberseite des Blattes orientiert, während das Phloem immer zur morphologischen Unterseite orientiert ist.
Die Mittelrippe ist seitlich mit kleineren Seitenvenen verbunden. Jeder von ihnen ist mit noch kleineren verbunden, und diese wiederum sind in noch kleinere unterteilt und so weiter. Die Anzahl der Verzweigungsreihenfolgen in Dicots variiert von 2 bis 5 oder mehr. Die kleinsten Äste bilden Zellen, in denen geschlossen sind kleine Flächen Mesophyll - Warzenhof.
Im offenen dichotomen Venensystem gibt es keine geschlossenen Zellen.
Bei einkeimblättrigen Blättern (mit paralleler Venation) sind die Längsadern durch kleinere Adern miteinander verbunden - Kommissurenbündel, die in Form einfacher Querbrücken angeordnet sind. Die Anzahl der leitfähigen Gewebe in kleinen Venen nimmt allmählich ab. An den Enden der Bündel erstrecken sich die Xylem-Elemente oft über eine größere Distanz als die Phloem-Elemente. Das Xylem an den Enden der Venen besteht normalerweise aus kurzen Trachealelementen, das Phloem aus kurzen schmalen Siebelementen und großen Satellitenzellen.
Kleine Gefäßbündel, die im Mesophyll lokalisiert sind, sind von einer oder mehreren Schichten kompakt gefalteter Zellen umgeben, die die Hülle des Bündels bilden. Die Bündelscheiden können parenchymal oder sklerenchymal sein. Bei einigen Arten wird eine Suberinisierung der Zellwände in den Hüllen der Faszikel beobachtet, was darauf hindeutet, dass die Hüllenzellen als Endoderm fungieren können.
Collenchym und Sklerenchym befinden sich in großen Venen auf einer oder beiden Seiten. Das mit großen Venen verbundene Gewebe erhebt sich über die Blattoberfläche und bildet Vorsprünge. Der Gewebebereich, der zwischen den Vorsprüngen der Venen eingeschlossen ist, wird als "Interkostalzone" bezeichnet (lat. Der Name des Vorsprungs ist "Costa" - eine Rippe).
Bei Arten mit dicken Blättern finden sich im Mesophyll verschiedene Skleriden, die als Abstandshalter fungieren.
Die Blätter von Getreide sind durch eine starke Entwicklung von Sklerenchym gekennzeichnet.
Die Blätter von Gymnospermen sind in ihrer Struktur weniger vielfältig als die Blätter von Angiospermen. Überlegen Sie am Beispiel von Kiefernnadeln, wie sich die Anpassungsfähigkeit an niedrige Temperaturen im Winter und die Fähigkeit, mehrere Jahre zu leben, in der anatomischen Struktur widerspiegeln. Die Nadel hat eine dickwandige Epidermis mit einer kräftigen Cuticula und tief eingetauchten Stomata. Die Seitenzellen einiger Koniferen (Cupressaceae) haben einen kutikulären Grat, der über den Schließzellen der Spaltöffnungen hängt. Die Spaltöffnungen sind in vertikalen Reihen auf allen Seiten der Nadeln angeordnet. Unter der Oberhaut befinden sich dickwandige, teilweise verholzte Zellmembranen der Unterhaut. Das Mesophyll ist gefaltet. Es enthält Harzpassagen. In der Mitte der Nadel befinden sich leitfähige Bündel. Meist zwei, selten einer. Diese Bündel sind durch einen sklerenchymalen Estrich verbunden. Die Bündel sind von Transfusionsgewebe umgeben, das aus Tracheiden und Parenchymzellen besteht. Transfusionsgewebe (lat. transfusio - Transfusion) steht im Zusammenhang mit dem Transport von Wasser und Nährstoffen zwischen den Leitbündeln und dem Mesophyll.
Das Transfusionsgewebe ist von dickwandigem Entoderm umgeben. Das Endoderm hat in den frühen Entwicklungsstadien oft casparische Bänder und in den späteren Stadien eine suberische Platte. Im reifen Zustand haben Endodermzellen sekundär verholzte Membranen.
Die anatomische Struktur der Blattspreite weist eine erhebliche Plastizität auf. Die Struktur ändert sich in Abhängigkeit von den Lebensbedingungen der Pflanze und der Anordnung der Blätter an der Pflanze.
Eine der Richtungen in der Entwicklung der modernen Botanik – die ökologische Anatomie der Pflanzen – haben wir bereits angemerkt und gesagt, dass es der lebende Inhalt der Zelle – der Protoplast – ist, der in erster Linie auf Veränderungen der Umweltbedingungen reagiert. Daher sind die Zellen nördlicher Pflanzen weniger vakuolisiert und haben große Chloroplasten. Es gibt viele Stärkekörner im Stroma. Das Thylakoidsystem von Chloroplasten ist weniger entwickelt als bei Pflanzen gemäßigter Breiten. Die Anzahl der Mitochondrien in nördlichen Pflanzen ist 2-2,5 mal höher. Der EPR und der Golgi-Apparat sind weiter entwickelt. Solche Merkmale der submikroskopischen Struktur der Zellen des Mesophylls nördlicher Pflanzen werden als wichtige Anpassungsmerkmale charakterisiert, die zum Überleben von Pflanzen unter rauen klimatischen Bedingungen beitragen.
Bei Pflanzen, die an Feuchtigkeitsmangel angepasst sind, werden kalte, dicke Zellmembranen festgestellt, insbesondere in der Epidermis. Die stark verdickten Außenmembranen der Großzell-Epidermis nehmen die Hälfte der Zellhöhle ein. Oberhalb der Epidermis befindet sich eine dicke Kutikulaschicht. Von der Oberfläche ist die Kutikula mit dicken Wachsschuppen bedeckt, die den bläulichen Farbton der Blätter bestimmen.
Die Stomata sind in Vertiefungen eingetaucht, die von epidermalen Haaren ausgestoßen werden.
Sukkulenten haben Wasserspeichergewebe.
Interessante Anpassungen zum Einfangen von Feuchtigkeit in den Blättern von weicher Quinoa (Afrika). Das Blatt von der Ober- und Unterseite ist mit einer durchgehenden Schicht aus mehrreihig angeordneten Bläschenhaaren bedeckt. Die Dicke dieser Schicht übersteigt die Dicke des Mesophylls deutlich und schützt lebendes Gewebe vor dem Austrocknen. Die Haare sind in der Lage, nachts Luftfeuchtigkeit einzufangen und die Härte zu verbessern Wasserregime Pflanzen.
Blätter und Stängel der Quinoa unserer weitläufigen Waldzone sind mit zahlreichen bläschenartigen Haaren bedeckt, die den Eindruck eines pulvrigen Überzugs erwecken, aber sie bilden keine so dichte mehrreihige Schicht wie bei der weichen Quinoa.
Am Beispiel des Vergleichs der Blätter zweier Quinoa-Arten ist ersichtlich, dass die systematischen Merkmale ähnlicher taxonomischer Einheiten, wenn sie unter unterschiedlichen ökologischen Bedingungen angesiedelt sind, eine ungleiche Entwicklung erhalten und anpassungsfähiger Natur sind.
Die Blätter von Regenwaldpflanzen (wie Monstera) haben perforierte Blattspreiten und lange Blattstiele. Überschüssiges Wasser läuft immer leicht ab. Die Stomata sind offen. Palisadengewebe mit großen Lufteinschlüssen.
Neben Temperatur und Luftfeuchtigkeit ist Licht ein weiterer wichtiger abiotischer Faktor, auf den die Pflanze mit strukturellen Veränderungen reagiert.
In direktem Sonnenlicht sich entwickelnde Blätter sind kleiner als Schattenblätter, aber dicker als Schattenblätter, was auf die starke Entwicklung des Palisadenmesophylls zurückzuführen ist.
Evolutionsbedingte Anpassungen an Umweltbedingungen beziehen sich also auf quantitative Unterschiede, ändern aber nicht die Organisationsprinzipien.
Blattontogenese
Die Blätter sind in Form kleiner Höcker als seitliche Auswüchse des apikalen Meristems angelegt. An der Bildung des Blatttuberkels ist nicht nur die Tunika beteiligt, sondern auch die Körperzellen. Die Blattanlagen wachsen vom anfänglichen Vorsprung nach oben in Form eines konischen oder nadelförmigen Auswuchses. Bald umgeben die Blattrudimente das apikale Meristem und schützen es sowohl mechanisch als auch durch die Wärme, die sie während der Atmung freisetzen.
Das frühe Blattwachstum wird normalerweise in apikal und marginal (marginal) unterteilt. Die erste ist mit einer Verlängerung der Anlagen verbunden, die zweite mit seitlichem Wachstum, was zur Bildung von zwei Teilen der Platte führt. Dementsprechend hat die wachsende Primordia ein Apikalmeristem an der Spitze und zwei Randmeristeme an den Seiten der Achse. Das apikale Wachstum der Primordia ist nicht lang. Dem Wachstum der Spitze und der Ränder der Platte folgt interkalares Wachstum.
In einem zusammengesetzten Blatt ist die Aktivität des Randmeristems in separaten Zentren lokalisiert, von denen jedes ein separates Blatt bildet.
Die Entwicklung der Blattspreite ist schneller als das Wachstum des Blattstiels, der später durch interkalares Wachstum gebildet wird.
Wenn an der Grenze zwischen der Basis der Interkalar- und Randzonen eine Einschnürung gebildet wird, entwickelt sich ein Blattstiel. Der größte Teil des Blattstiels wird von der Interkalarzone gebildet. Wenn eine solche Einschnürung (an der Grenze zwischen der Basis der Interkalar- und Randzonen) nicht gebildet wird, entwickelt sich ein ungestieltes Blatt.
Im Blattrudiment einkeimblättriger Pflanzen werden kaum Randmeristeme gebildet, und interkalares Wachstum ist für lange Zeit charakteristisch. Daher ist die Hauptform der Blattspreite linear.
Unterschiede in der Teilungsrate und Dehnung von Zellen in verschiedenen Schichten der Platte führen zur Ausbildung zahlreicher Interzellularräume und einer typischen Mesophyllstruktur.
Die Entwicklung anderer Gewebe korreliert mit der Entwicklung leitfähiger Gewebe. Das Procambium trennt sich gleichzeitig im Blatt und im angrenzenden Bereich des Wachstumskegels und bildet einen durchgehenden Strang. Es bildet das primäre Leitbündel, das kontinuierlich und dem Blatt und dem Stängel gemeinsam ist.
Die Ontogenese des Blattes zeugt von der gemeinsamen Natur des Blattes und des Stängels als Teile eines einzigen Ganzen – des Sprosses.
Von dem Moment an, in dem sich die Knospe entwickelt, beginnt die Extra-Knospen-Phase der Blattentwicklung. Gleichzeitig vergrößert sich die Oberfläche der Blätter um das Hundert- bis Tausendfache. Das Oberflächenwachstum wird erreicht, indem die meisten Blattzellen geteilt und in Länge und Breite gestreckt werden.
Die Lebensdauer der Blätter hängt von genetischen und klimatischen Faktoren ab. Bei Laubbäumen und Sträuchern gemäßigten Klimas beträgt die Extrabud-Zeit der Blattlebensdauer 4-5 Monate. Die Blätter von Nadelpflanzen leben zwei bis sechs oder mehr Jahre. Aber in jedem Fall ist die Lebensdauer der Blätter viel geringer als die Lebensdauer der axialen Organe der Pflanze.
Welwitschia mirabilis stammt aus den Steinwüsten im Südwesten Afrikas. Dieser Zwergbaum hat eine lange Wurzel, einen dicken und kurzen Stamm (bis zu 50 cm hoch und bis zu 1 m dick) und zwei große Blätter, die ein Leben lang bestehen bleiben (bis zu 2000 Jahre). Die Blätter erreichen eine Länge von 2-3 Metern und wachsen an der Basis ständig und sterben oben ab. Fast die einzige Feuchtigkeitsquelle für Velvichia ist dichter Nebel, dessen Feuchtigkeit diese Pflanze durch zahlreiche Spaltöffnungen auf beiden Seiten des Blattes aufnimmt (22.200 Spaltöffnungen pro 1 cm 2 ).
Die aktive Trennung von Blättern von Zweigen, ohne das lebende Gewebe zu beschädigen, wird als Abszision bezeichnet. Der saisonale Laubfall von Bäumen ist das Ergebnis einer Reaktion auf eine Änderung der Tageslänge. Fallende Blätter im Herbst reduzieren die Verdunstungsfläche der Pflanze erheblich, was bei physiologischem Feuchtigkeitsmangel im Herbst und Winter erforderlich ist. Dank des Laubfalls wird die Gefahr des Abbrechens von Ästen durch die Schneelast verringert.
Bei den meisten Blättern erfolgt die Bildung einer Trennschicht während der Ontogenese. In dieser Zone ist die Menge an mechanischem Gewebe reduziert. Thyle werden in den Trachealelementen vor dem Ablösen gebildet, und Callose wird in den Siebelementen abgelagert. Während der Abszision kommt es zu einer enzymatischen Zerstörung der Zellmembranen, was zur Trennung der Zellen voneinander führt. Zu den Veränderungen der Membranen gehören der Verlust der Zementierungskapazität der medianen Lamina (teilweise aufgrund der Entfernung von Kalzium aus ihr), die Hydrolyse der Zellulosemembranen selbst und das Reißen von sklerifizierten Trachealelementen.
Durch die Ablagerung von Schutzsubstanzen wie Suberin oder Wundgummi in den Membranen und Interzellularräumen entsteht eine Schutzschicht oder Narbe. Bei Holzgewächse die Schutzschicht wird durch das Periderm ersetzt, das sich unter der Schutzschicht niederlegt und mit dem Periderm des restlichen Stiels kontinuierlich wird.
Das Fallenlassen von Blättern ist nicht unbedingt mit dem Prozess der Auflösung von Zellmembranen verbunden. Beim Fallenlassen von Fichtennadeln kann eine mechanische Ablösung ohne chemische Vorveränderung beobachtet werden.
Nach dem Laubfall bleiben Blattnarben mit ihren Büschelnarben zurück. Balkennarben- Dies sind die gebrochenen Enden der Leitbündel, die von den Blattspuren zum Blattstiel gingen, bevor es abfiel.
Blattspuren- Dies sind Äste vom Leitungssystem des Stängels zu den Blättern. Die Blattspur erstreckt sich von der Stelle ihres Zusammenflusses mit dem Stammbündel bis zum Austritt in das Blatt. Ein Blatt kann eine oder mehrere Blattspuren aufweisen.
Da Stamm und Blatt haben einen phylogenetisch gemeinsamen Ursprung, es gibt keinen grundsätzlichen Unterschied zwischen Blattspuren und Stammbündeln. Die entsprechenden Begriffe haben eine beschreibende, topografische Bedeutung.
Im Bereich des Knotens, wo die Blattspur vom zentralen Leitzylinder des Stiels zur Blattbasis abweicht, bildet sich im Zylinder eine Parenchymzone - eine Blattlücke oder eine Blattlücke. Die Anzahl der Blattspuren und Blattlücken variiert von Pflanze zu Pflanze.
Der primäre Knotentyp bei Angiospermen ist einer mit drei oder mehr Lücken. Die Hauptentwicklungsrichtung der Angiospermen ist durch eine Abnahme der Anzahl von Lakunen gekennzeichnet.
Die Lakunen, das Mark, die interfaszikulären Zonen, das Leitungssystem und das an der Peripherie des Leitungssystems befindliche Pericyclus bilden den inneren Kern des axialen Teils der Pflanze (Stamm und Wurzel) oder Stele.
Die Klassifizierung der Stelen basiert auf der gegenseitigen Anordnung von leitfähigen und nicht leitfähigen Geweben in den Achsenorganen im Primärstadium der Entwicklung. Beim einfachsten Stelentyp, der als der phylogenetisch primitivste gilt, bildet das leitfähige Gewebe eine durchgehende Säule, wobei das Phloem das Xylem umgibt. Das ist eine Protostele. Eine solche Organisation des Stammes ist biologisch unvollkommen, weil die Kontaktfläche zwischen leitfähigem und darunterliegendem Gewebe ist klein.
Die weitere Entwicklung der Stele folgte dem Weg, die Kontaktfläche der leitfähigen Gewebe mit den Hauptgeweben zu vergrößern. Dies wurde auf zwei Wegen erreicht. In einem Fall bildet die Stele tiefe Auswüchse zur Rinde hin und hat im Querschnitt die Form eines Sterns. Das ist eine Aktinostele.
Der zweite, biologisch vielversprechendere Entwicklungsweg ist das Auftreten eines Parenchymkerns im Zentrum der Stele. Das ist ein Siphon.
Mit der Entwicklung der Blätter entsteht eine engere Verbindung zwischen Leit- und Grundgewebe. Je mehr Blätter, desto mehr ist die Stele mit dem Hauptparenchym gesättigt. Die Stele nimmt die Form eines Netzes (griech. Netz - Diktat) an. Dies ist eine Diktiostele.
Die Stele moderner Blütenpflanzen ist noch parenchymatisierter. Die Stele, ein System aus leitfähigen Strängen und interfaszikulären Zonen, ist Eustela (griechisch eu - gut).
Bei Monokotylen erreichen Blattspuren die Mitte des Stängels und weichen dann zur Peripherie ab. Daher scheinen die Bündel auf dem Querschnitt des Stammes ohne jede Ordnung verstreut zu sein. Ataktos aus dem Griechischen. chaotisch. Daher der Name der Stele von Monocots - Ataktostele. Sie ist charakterisiert hochgradig Parenchymisierung, wenn jedes Blatt Spur "
Die Wurzel ist ein axiales Organ, das radialsymmetrisch ist und in der Länge zunimmt, solange das apikale Meristem erhalten bleibt. Die Wurzel unterscheidet sich morphologisch vom Stängel dadurch, dass Blätter niemals darauf erscheinen und das apikale Meristem von einer Wurzelkappe bedeckt ist. Die Verzweigung und Initiierung von Adventivknospen in Wurzelnachkommenpflanzen erfolgt endogen (intern) als Ergebnis der Aktivität des Pericyclus (primäres laterales Meristem).
Root-Funktionen:
1) Die Wurzel nimmt Wasser aus dem Boden mit darin gelösten Mineralien auf;
2) spielt eine Ankerrolle und fixiert die Pflanze im Boden;
3) dient als Behälter für Nährstoffe;
4) nimmt an der Primärsynthese bestimmter organischer Substanzen teil;
5) in Wurzelnachkommenpflanzen erfüllt es die Funktion der vegetativen Reproduktion.
Root-Klassifizierung. Nach Herkunft werden die Wurzeln in Haupt-, Adventiv- und Seitenwurzeln unterteilt. Die Wurzel, die sich aus der Keimwurzel des Samens entwickelt, wird genannt hauptsächlich; Wurzeln, die an anderen Pflanzenorganen (Stamm, Blatt, Blüte) entstehen, werden genannt Adnex. Die Rolle der Adventivwurzeln im Leben der krautigen Angiospermen ist enorm, da das Wurzelsystem in erwachsenen Pflanzen (sowohl Monokotyledonen als auch viele Dikotyledonen) hauptsächlich (oder nur) aus Adventivwurzeln besteht. Das Vorhandensein von Adventivwurzeln auf dem basalen Teil der Triebe macht es einfach, Pflanzen künstlich zu vermehren, indem man sie in separate Triebe oder Gruppen von Trieben mit Adventivwurzeln teilt.
In jungen Stämmen werden die Rudimente von Adventivwurzeln endogen gebildet - aus Zellen des interfaszikulären Parenchyms, in alten - aus dem Parenchym in der Nähe des Kambiums der Kernstrahlen, dh dem Kambium parenchymalen Ursprungs. Zufällige Wurzeln monokotyler Stängel entstehen im Parenchym, das die Bündel umgibt.
Seite Wurzeln werden an den Haupt- und Nebenwurzeln gebildet. Durch ihre weitere Verzweigung entstehen Seitenwurzeln höherer Ordnung. Am häufigsten erfolgt die Verzweigung bis zur vierten oder fünften Ordnung.
Die Gesamtheit aller Wurzeln einer Pflanze wird als bezeichnet Wurzelsystem.
nach Herkunft:
Hauptwurzelsystem entwickelt sich aus der Keimwurzel und wird durch die Hauptwurzel (erster Ordnung) mit Seitenwurzeln zweiter und nachfolgender Ordnung repräsentiert. Bei vielen Bäumen und Sträuchern sowie bei einjährigen und einigen mehrjährigen krautigen Zweikeimblättrigen entwickelt sich nur das Hauptwurzelsystem;
Adventives Wurzelsystem entwickelt sich auf Stängeln, Blättern, manchmal auf Blüten. Dieser Wurzelursprung wird als primitiver angesehen, da er für höhere Sporen charakteristisch ist, die nur ein System von Adventivwurzeln haben. Das System der Adventivwurzeln in Angiospermen wird aus dem Samen gebildet, in dem sich das Protokorm (Embryonalknolle) entwickelt, und anschließend - Adventivwurzeln darauf;
gemischtes Wurzelsystem weit verbreitet unter zweikeimblättrigen und einkeimblättrigen Pflanzen. Bei einer Pflanze, die aus einem Samen gezogen wird, entwickelt sich zuerst das System der Hauptwurzel, aber sein Wachstum hält nicht lange an - es hört oft im Herbst der ersten Vegetationsperiode auf. Zu diesem Zeitpunkt entwickelt sich konsequent ein System von Adventivwurzeln auf dem Hypokotyl, Epikotyl und den nachfolgenden Metameren des Haupttriebs und anschließend auf dem basalen Teil der Seitentriebe. Je nach Pflanzenart werden sie in bestimmten Teilen von Metameren (an Knoten, unter und über Knoten, an Internodien) oder über ihre gesamte Länge initiiert und entwickelt.
Klassifizierung von Wurzelsystemen informieren.
Das Hauptwurzelsystem wird aufgerufen ausschlaggebend wenn die Hauptwurzel die Seitenwurzeln in Länge und Dicke merklich übersteigt;
Bei ähnlicher Größe der Haupt- und Seitenwurzeln entsteht das Wurzelsystem faserig. Ein gemischtes Wurzelsystem kann auch sein ausschlaggebend, wenn die Hauptwurzel viel größer ist als die anderen, faserig wenn alle Wurzeln relativ gleich groß sind. Die gleichen Begriffe gelten für das System der Adventivwurzeln.
Innerhalb desselben Wurzelsystems führen Wurzeln oft unterschiedliche Funktionen aus. Es gibt Skelettwurzeln (stützend, stark, mit entwickelten mechanischen Geweben), Wachstumswurzeln (schnell wachsend, aber wenig verzweigend), Saugwurzeln (dünn, kurzlebig, intensiv verzweigend).



































- Größe: 2 MB
- Anzahl der Folien: 36
Beschreibung der Präsentation WURZEL UND WURZELSYSTEME 1. Funktionen und evolutionäre Folien
WURZEL UND WURZELSYSTEME 1. Funktionen und evolutionärer Ursprung der Wurzel. 2. Die Primärstruktur der Wurzel. 3. Sekundäre Wurzeländerungen. 4. Bildung von Seiten- und Adventivwurzeln. Wurzelsysteme. 5. Spezialisierung und Modifikation der Wurzeln.
Die Wurzel ist ein axiales Organ, das radialsymmetrisch ist und aufgrund der Aktivität des apikalen Meristems unendlich lang wird. Blätter erscheinen nie an der Wurzel, und das apikale Meristem ist immer mit einer Kappe bedeckt. Die Hauptfunktion der Wurzel ist die Aufnahme von Wasser und Mineralien, dh die Bereitstellung von Bodennahrung für die Pflanze. Neben der oben genannten Hauptfunktion erfüllen die Wurzeln noch weitere Funktionen: Sie stärken die Pflanze im Boden, ermöglichen das vertikale Wachstum und den Auftrieb; in den Wurzeln gibt es eine Sekundärsynthese verschiedener Substanzen (Aminosäuren, Alkaloide, Phytohormone usw.); Ersatzsubstanzen können in den Wurzeln abgelagert werden; Wurzeln interagieren mit den Wurzeln anderer Pflanzen, Bodenmikroorganismen und Pilzen.
Wurzeln entstanden aus den Telomen von Rhinophyten, die sich über die Erdoberfläche ausbreiteten. Im Laufe der Evolution begannen einige Zweige dieser Telome, sich in den Boden zu vertiefen und Wurzeln zu bilden.
Die Wurzeln sind an eine perfektere Bodenernährung angepasst. Die Entstehung der Wurzeln war begleitet von einer tiefgreifenden Umstrukturierung ihrer gesamten Struktur. Sie haben spezialisierte Gewebe. Die Funktion, Substanzen aus dem Boden aufzunehmen, wurde von jungen Wurzelenden übernommen. Sie halten lebende Zellen an der Oberfläche. Diese Zellen bildeten das funktionell wichtigste Wurzelgewebe, das Rhizoderm.
Die Funktion, Substanzen aus dem Boden aufzunehmen, wurde von jungen Wurzelenden übernommen. Sie halten lebende Zellen an der Oberfläche. Diese Zellen bildeten das funktionell wichtigste Wurzelgewebe, das Rhizoderm. Darüber hinaus nahm im Laufe der Evolution die absorbierende Wurzeloberfläche aufgrund von drei Faktoren zu: 1) reichliche Verzweigung und die Bildung einer großen Anzahl von absorbierenden Enden; 2) das ständige Wachstum von Wurzeln und die Bewegung von Saugenden zu neuen Bodenbereichen; 3) die Bildung von Wurzelhaaren.
Da das Wurzelwachstum in dichtem Boden stattfindet, muss sein apikales Meristem geschützt werden. Das apikale Meristem wurde durch das Auftreten einer Wurzelkappe vor Beschädigung geschützt. Das Auftreten von Wurzeln wurde durch eine Zunahme der Trockenheit des Klimas verursacht. Das Einsetzen eines trockeneren Klimas hat dazu geführt, dass sich Landpflanzen an das Substrat anheften und Wasser und Nährstoffe daraus aufnehmen. Allerdings hat sich im Laufe der Evolution die Struktur der Wurzel bei verschiedenen Pflanzenarten weniger verändert als die des Stängels. Dies liegt daran, dass die Bedingungen im Bodenmilieu stabiler sind als in der Luft. Daher gilt die Wurzel als ein "konservativeres" Organ, obwohl sie viel später als die Flucht auftauchte. Die Wurzelbildung ist eine wichtige Pflanzenaromorphose. Dank ihm konnten die Pflanzen trockenere Böden meistern und große aufstrebende Triebe bilden.
Reaktion von Amyloplasten der Wurzelkappe auf die Schwerkraft. Verschieben von Statolithen spielt wichtige Rolle bei der Schaffung von Phytohormongradienten, die für vertikales Wurzelwachstum sorgen.
Die Struktur der Wurzel eines Weizensämlings (Triticum aestivum): A - Diagramm der Wurzelstruktur; B — Differenzierung von Rhizoderm- und Exodermzellen. 1 - Leitungszone, 2 - Saugzone, 3 - Dehnungszone, 4 - Teilungszone, 5 - Wurzelhaar, 6 - Wurzelkappe.
Querschnitt der Wurzel (a - einkeimblättrige, b - zweikeimblättrige Pflanze)
Der primäre Cortex entsteht aus dem Periblem. Seine Hauptmasse besteht aus lebenden Parenchymzellen mit dünnen Membranen. Zwischen ihnen bildet sich ein System von Interzellularräumen, die entlang der Wurzelachse verlängert sind. Gase (CO 2) zirkulieren durch die Interzellularräume. Gase sind notwendig, um einen intensiven Stoffwechsel in den Zellen der Rinde und der Rhizodermis aufrechtzuerhalten. Ein energetischer Stoffwechsel in den Zellen der Rinde ist notwendig, um eine Reihe wichtiger Funktionen zu erfüllen: 1) Die Zellen der Rinde versorgen das Rhizoderm mit plastischen Substanzen und sind selbst an der Aufnahme und Weiterleitung von Substanzen beteiligt; 2) verschiedene Substanzen werden im Kortex synthetisiert, die dann auf andere Gewebe übertragen werden; 3) Reservesubstanzen reichern sich in den Zellen der Hirnrinde an; 4) In der Rinde befinden sich oft Pilzhyphen, die Mykorrhiza bilden.
Endodermzellen durchlaufen drei Entwicklungsstadien. In der Absorptionszone befindet sich das Endoderm im ersten Stadium. In der Mitte der radialen Wände seiner Zellen bilden sich casparianische Bänder. Casparian-Bands blockieren die Bewegung von Substanzen durch die Zellmembranen, dh entlang des Apoplasten. Das zweite Stadium kann in der Zone der Seitenwurzeln beobachtet werden. Gleichzeitig erscheint auf der Innenseite der Zellmembran eine dünne Suberinplatte. Das Endoderm ist jedoch immer noch frei, Lösungen zu passieren, da es einzelne Zellen mit dünnen Wänden enthält. Das dritte Entwicklungsstadium des Endoderms ist im Bereich der Wurzeln von Monokotylen zu beobachten. Die inneren und radialen Wände seiner Zellen sind stark verdickt. Auf Querschnitten haben diese Zellen eine Hufeisenform. Es gibt keine Kontrollpunkte. Das dickwandige Endoderm schützt leitfähiges Gewebe und erhöht die Wurzelfestigkeit.
Der Casparian-Gürtel ist eine wasserdichte Barriere, die Wasser zwingt, den Apoplasten zu verlassen und durch die Membranen der Endodermzellen in den Symplasten zu strömen.
Wasserzufluss vom Boden zur Wurzel: Wasser kann sich entlang des Apoplasten und Symplasten bewegen, bis es die Endodermis erreicht. Eine weitere Bewegung entlang des Apoplasten ist nicht möglich.
Verschiedene Strukturtypen des Zentralzylinders der Wurzel (Primärstruktur): A-Diarchie, B-Triarchie, C-Tetrarchie, G-Polyarchie. Typen A-B charakteristisch für Dikotyledonen, G - in vielen Monokotyledonen. 1 - Bereich des primären Kortex, 2 - primäres Phloem, 3 - primäres Xylem.
Es ist möglich, 4 Stadien des Übergangs der Wurzel von der Primär- zur Sekundärstruktur zu unterscheiden: 1) das Auftreten des Kambiums zwischen den Bereichen des primären Phloems und des Xylems; 2) Bildung von Phellogen durch das Pericyclus; 3) Ablösung der primären Kruste; 4) Änderung der radialen Anordnung von leitenden Geweben durch kollaterale.
Übergang von der primären zur sekundären Wurzelstruktur , 11 - sekundäre Rinde, 12 - Phellogen, 13 - Phellem.
Schema der primären Differenzierung von leitfähigen Geweben in der Erbsenwurzel 1 - zentripetale Differenzierung von Xylem, 2 - Epidermis, 3 - primärer Kortex, 4 - Endodermis, 5 - erste differenzierte Xylemelemente, 6 - undifferenzierte Xylemelemente, 7 - erste differenzierte Phloemelemente , 9 - apikales Meristem, 10-Wurzel-Kappe.
Sekundäre Veränderungen in der Wurzel von Monokotylen. Die überwiegende Mehrheit der einkeimblättrigen Pflanzen behält die primäre Wurzelstruktur bis zum Ende ihres Lebens. Viele Elemente der Wurzel unterliegen jedoch einer Verholzung. Bei verholzten Monokotylen (Palmen, Dracaena, Yucca) entsteht eine Meristemschicht in der Wurzelrinde aus Parenchymzellen oder aus dem Perizykel. Daraus werden Reihen geschlossener Leiterbündel gebildet. Nach dieser Reihe von Leitungsbündeln erscheint eine neue Schicht von Bildungsgewebe im peripheren Teil des Parenchyms der primären Rinde. Aus dieser Meristemschicht entsteht eine neue Reihe von Leitbündeln. Dadurch verdickt sich die Wurzel.
Adventivwurzeln entstehen an verschiedenen Pflanzenorganen - an Stängeln, Blättern und Wurzeln. Adventivwurzeln, die am Stamm entstanden sind, heißen Stammwurzeln, solche, die an der Wurzel entstanden sind, heißen Wurzelwurzeln. Seiten- und Adventivwurzeln sind endogenen Ursprungs, dh sie werden in das innere Gewebe gelegt.
Die laterale Wurzelinitiation beginnt mit der Zellteilung des Perizykels. In diesem Fall bildet sich auf der Oberfläche der Stele ein meristematischer Tuberkel. Nach einer Reihe von Zellteilungen des meristematischen Tuberkels erscheint eine Seitenwurzel. Es hat ein eigenes apikales Meristem und eine Scheide. Die Knospe der Seitenwurzel wächst, durchbricht die Primärrinde der Mutterwurzel und bewegt sich nach außen. Normalerweise entstehen Seitenwurzeln gegen Xylemelemente. Daher sind sie in regelmäßigen Längsreihen entlang der Wurzel angeordnet. Sie treten in der Absorptionszone oder etwas höher auf. Die Verlegung der Seitenwurzeln erfolgt akropetal, d.h. von der Wurzelbasis bis zur Wurzelspitze.
Adventive Wurzeln werden normalerweise in Geweben gelegt, die zur meristematischen Aktivität fähig sind: im Pericyclus, Kambium, Phellogen. Die endogene Bildung von seitlichen (und zufälligen) Wurzeln hat einen adaptiven Wert. Wenn die Verzweigung in der Spitze stattfinden würde, wäre das Vordringen der Wurzel im Boden schwierig.
Dichotome Verzweigung im Wurzelsystem von Bärlapp (Lycopodium clavatum) 1 - isotomische dichotome Verzweigung der dünnsten Wurzeln
Mykorrhiza: A - ektotrophe Eichenmykorrhiza, B, C - endotrophe Orchideenmykorrhiza. Knötchen an Lupinenwurzeln