Šoninių šaknų formavimasis. Šaknis, jos funkcijos. Šaknų tipai ir šaknų sistemos
Šaknis yra požeminis ašinis augalų elementas, kuris yra svarbiausia jų dalis, pagrindinis vegetatyvinis organas. Dėl šaknies augalas fiksuojamas dirvožemyje ir laikomas ten visą savo gyvavimo ciklą, taip pat aprūpinamas joje esančiu vandeniu, mineralais ir maistinėmis medžiagomis. Yra įvairių šaknų rūšių ir tipų. Kiekvienas iš jų turi savo išskirtines savybes. Šiame straipsnyje mes apsvarstysime esamus šaknų tipus, šaknų sistemų tipus. Taip pat susipažinsime su jiems būdingais bruožais.
Kokios yra šaknų rūšys?
Standartinė šaknis būdinga siūliška arba siaura cilindro forma. Daugelyje augalų, be pagrindinės (pagrindinės) šaknies, išsivysto ir kitų rūšių šaknys – šoninės ir atsitiktinės. Pažvelkime atidžiau, kas jie yra.
pagrindinė šaknis
Šis augalo organas išsivysto iš sėklos gemalinės šaknies. Visada yra viena pagrindinė šaknis (kitų rūšių augalų šaknys dažniausiai būna daugiskaitos). Jis išlieka augale visą gyvavimo ciklą.
Šaknims būdingas teigiamas geotropizmas, tai yra, dėl gravitacijos ji gilėja į substratą vertikaliai žemyn.
atsitiktinės šaknys
Atsitiktiniais vadinamos augalų šaknų rūšys, kurios susidaro ant kitų jų organų. Šie organai gali būti stiebai, lapai, ūgliai ir kt. Pavyzdžiui, javai turi vadinamąsias pirmines atsitiktines šaknis, kurios yra įterptos į sėklos gemalų kotelį. Jie vystosi sėklų dygimo procese beveik kartu su pagrindine šaknimi.
Taip pat yra lapinių atsitiktinių šaknų tipų (susidaro dėl lapų įsišaknijimo), stiebo arba mazginių (susidaro iš šakniastiebių, antžeminių ar požeminių stiebo mazgų) ir kt. Apatiniuose mazguose susidaro galingos šaknys, kurios vadinamos antenos (arba atraminės).
Atsitiktinių šaknų atsiradimas lemia augalo gebėjimą vegetatyviškai daugintis.
Šoninės šaknys
Šoninės yra vadinamos šaknimis, kurios atsiranda kaip šoninė šaka. Jie gali susidaryti tiek ant pagrindinių, tiek ant atsitiktinių šaknų. Be to, jie gali atsišakoti nuo šoninių, dėl to susidaro aukštesnės eilės (pirmos, antros ir trečios) šoninės šaknys.
Dideliems šoniniams organams būdingas skersinis geotropizmas, tai yra, jų augimas vyksta beveik horizontalioje padėtyje arba kampu į dirvos paviršių.
Kas yra šaknų sistema?
Šaknų sistema vadinama visų rūšių ir tipų šaknys, kurias turi vienas augalas (tai yra, jų visuma). Priklausomai nuo pagrindinių, šoninių ir papildomų šaknų augimo santykio, nustatomas jo tipas ir pobūdis.
Šaknų sistemų tipai
Jei pagrindinė šaknis yra labai gerai išvystyta ir pastebima tarp kitos rūšies šaknų, tai reiškia, kad augalas turi lazdelių sistemą. Daugiausia randama dviskilčiuose augaluose.
Šio tipo šaknų sistemai būdingas gilus daigumas į dirvą. Taigi, pavyzdžiui, kai kurių žolių šaknys gali prasiskverbti į 10–12 metrų gylį (erškėtis, liucerna). Medžių šaknų įsiskverbimo gylis kai kuriais atvejais gali siekti 20 m.
Jei papildomos šaknys yra ryškesnės, dauginasi, o pagrindinė pasižymi lėtu augimu, tada susidaro šaknų sistema, vadinama pluoštine.

Paprastai tokia sistema būdinga ir kai kuriems žoliniams augalams. Nepaisant to, kad pluoštinės sistemos šaknys neįsiskverbia taip giliai kaip strypų sistemos, jos geriau supina greta jų esančias dirvožemio daleles. Daug palaidų krūmų ir šakniastiebių žolių, formuojančių gausias pluoštines plonas šaknis, plačiai naudojamos dauboms, šlaitų dirvožemiams sutvarkyti ir kt. Geriausios velėnos žolės yra belapės laužos, eraičinai ir kt.

modifikuotos šaknys
Be tipiškų aukščiau aprašytų, yra ir kitų rūšių šaknų ir šaknų sistemų. Jie vadinami modifikuotais.
saugojimo šaknys
Atsargos apima šakniavaisius ir šakniagumbius.
Šakniavaisiai yra pagrindinės šaknies sustorėjimas dėl maistinių medžiagų nusėdimo joje. Taip pat apatinė stiebo dalis dalyvauja formuojant šakniavaisį. Jį daugiausia sudaro laikymo bazinis audinys. Šakniavaisių pavyzdžiai yra petražolės, ridikai, morkos, burokėliai ir kt.

Jei sustorėjusios saugojimo šaknys yra šoninės ir atsitiktinės šaknys, tada jos vadinamos šakniagumbiais (spurgais). Jie išsivysto bulvėse, saldžiosiose bulvėse, jurginuose ir kt.
oro šaknys
Tai šoninės šaknys, augančios oro dalyje. Pateikti numeriu atogrąžų augalai. Vanduo ir deguonis yra absorbuojami iš oro. Galima naudoti tropiniuose augaluose, augančiuose trūkumo sąlygomis mineralai.

kvėpavimo šaknys
Tai savotiškos šoninės šaknys, kurios auga aukštyn, kyla virš substrato paviršiaus, vandens. Tokio tipo šaknys susidaro augaluose, augančiuose per drėgnose dirvose, pelkių sąlygomis. Tokių šaknų pagalba augmenija trūkstamą deguonį gauna iš oro.
Atraminės (lentos formos) šaknys
Tokio tipo medžių šaknys būdingos stambioms rūšims (bukai, guobos, tuopos, atogrąžų ir kt.) Tai trikampės vertikalios ataugos, suformuotos iš šoninių šaknų ir einančios šalia arba virš dirvos paviršiaus. Jos dar vadinamos lentos formos, nes primena lentas, kurios atsiremia į medį.
Atžalų šaknys (haustorija)
Tai papildomos atsitiktinės šaknys, besivystančios ant vijoklinių augalų stiebo. Jų pagalba augalai turi galimybę prisitvirtinti prie tam tikros atramos ir lipti (austyti) aukštyn. Tokios šaknys yra, pavyzdžiui, atkakliuose fikusuose, gebenėse ir kt.

Ištraukiamos (susitraukiančios) šaknys
Būdinga augalams, kurių šaknis ties pagrindu išilgine kryptimi smarkiai sumažinta. Pavyzdys galėtų būti augalai, turintys svogūnėlių. Ištraukiamos šaknys suteikia svogūnėliams ir šakniavaisiams tam tikrą įdubimą dirvoje. Be to, jų buvimas lemia tvirtą rozečių (pavyzdžiui, kiaulpienių) prigludimą prie žemės, taip pat vertikalaus šakniastiebio ir šaknies kaklelio požeminę padėtį.
mikorizė (grybelio šaknis)
Mikorizė yra aukštesniųjų augalų šaknų simbiozė (abipusiai naudingas sugyvenimas) su grybų hifais, kurie jas pina, veikdami kaip šaknų plaukeliai. Grybai aprūpina augalus vandeniu ir jame ištirpusiomis maistinėmis medžiagomis. Savo ruožtu augalai aprūpina grybus organinėmis medžiagomis, reikalingomis jų gyvybinei veiklai.
Mikorizė būdinga daugelio aukštesnių augalų, ypač sumedėjusių, šaknims.
bakteriniai mazgeliai
Tai modifikuotos šoninės šaknys, pritaikytos simbiotiniam gyvenimui su azotą fiksuojančiomis bakterijomis. Mazgelių susidarymas atsiranda dėl jaunų šaknų įsiskverbimo į vidų. Toks abipusiai naudingas bendras gyvenimas leidžia augalams gauti azotą, kurį bakterijos iš oro perneša į jiems prieinamą formą. Kita vertus, bakterijoms suteikiama speciali buveinė, kurioje jos gali veikti nekonkuruodamos su kitų tipų bakterijomis. Be to, jie naudoja medžiagas, esančias augalijos šaknyse.

Bakteriniai mazgeliai būdingi ankštinių šeimos augalams, kurie plačiai naudojami kaip meliorantai sėjomainose, siekiant praturtinti dirvas azotu. Geriausiais azotą kaupiančiais augalais laikomi liemeniniai ankštiniai augalai, tokie kaip mėlynoji ir geltonoji liucerna, raudonasis ir ašparnis, raguotas skėris ir kt.
Be minėtų metamorfozių, yra ir kitų rūšių šaknų, pavyzdžiui, atraminės šaknys (padeda sutvirtinti stiebą), dygliuotos šaknys (padeda augalams nenuskęsti skystame purve) ir šaknų čiulptukai (turi papildomus pumpurus ir užtikrina vegetatyvinį dauginimąsi).
Filogenetiškai šaknis atsirado vėliau nei stiebas ir lapas – tai susiję su augalų perėjimu į gyvybę sausumoje ir tikriausiai kilo iš į šaknis panašių požeminių šakų. Šaknis neturi nei lapų, nei pumpurų, išsidėsčiusių tam tikra tvarka. Pasižymi viršūniniu ilgiu, jo šoninės šakos kyla iš vidinių audinių, augimo vieta padengta šaknies kepurėle. Šaknų sistema formuojasi visą augalo organizmo gyvenimą. Kartais šaknis gali tarnauti kaip maistinių medžiagų nusėdimo vieta. Šiuo atveju jis yra modifikuotas.
Šaknų tipai
Pagrindinė šaknis susidaro iš gemalinės šaknies sėklų dygimo metu. Jis turi šonines šaknis.
Atsitiktinės šaknys išsivysto ant stiebų ir lapų.
Šoninės šaknys yra bet kokių šaknų šakos.
Kiekviena šaknis (pagrindinė, šoninė, atsitiktinė) turi galimybę šakotis, o tai žymiai padidina šaknų sistemos paviršių, o tai prisideda prie geresnio augalo sutvirtinimo dirvoje ir pagerina jo mitybą.
Šaknų sistemų tipai
Yra du pagrindiniai šaknų sistemų tipai: liemeninė šaknis, kurios pagrindinė šaknis yra gerai išvystyta, ir pluoštinė. Pluoštinė šaknų sistema susideda iš daugybės atsitiktinių, tokio paties dydžio šaknų. Visa šaknų masė susideda iš šoninių arba atsitiktinių šaknų ir atrodo kaip skiltis.
Labai išsišakojusi šaknų sistema sudaro didžiulį sugeriantį paviršių. Pavyzdžiui,
- bendras žieminių rugių šaknų ilgis siekia 600 km;
- šaknų plaukelių ilgis - 10 000 km;
- bendras šaknų plotas 200 m 2.
Tai daug kartų didesnis už antžeminės masės plotą.

Jei augalas turi aiškiai apibrėžtą pagrindinę šaknį ir išsivysto papildomos šaknys, tada susidaro mišri šaknų sistema (kopūstų, pomidorų).
Išorinė šaknies struktūra. Vidinė šaknies struktūra
Šaknų zonos

šaknies dangtelis
Šaknis auga į ilgį su savo galiuku, kur yra jaunos lavinamojo audinio ląstelės. Auganti dalis yra padengta šaknies kepurėle, kuri apsaugo šaknies galiuką nuo pažeidimų ir palengvina šaknies judėjimą dirvoje augimo metu. Pastaroji funkcija atliekama dėl šaknies kepurėlės išorinių sienelių savybės pasidengti gleivėmis, o tai sumažina trintį tarp šaknies ir dirvožemio dalelių. Jie netgi gali išstumti dirvožemio daleles. Šaknies kepurėlės ląstelės yra gyvos, dažnai jose yra krakmolo grūdelių. Dangtelio ląstelės nuolat atnaujinamos dėl dalijimosi. Dalyvauja teigiamose geotropinėse reakcijose (šaknų augimo kryptis į Žemės centrą).

Dalijimosi zonos ląstelės aktyviai dalijasi, šios zonos ilgis skiriasi skirtingose rūšyse ir skirtingose to paties augalo šaknyse.
Už padalijimo zonos yra išplėtimo zona (augimo zona). Šios zonos ilgis neviršija kelių milimetrų.
Pasibaigus linijiniam augimui, prasideda trečiasis šaknų formavimosi etapas - jos diferenciacija, formuojasi ląstelių diferenciacijos ir specializacijos zona (arba šaknų plaukelių ir absorbcijos zona). Šioje zonoje jau išskiriamas išorinis epiblemos sluoksnis (rizodermas) su šaknų plaukeliais, pirminės žievės sluoksnis ir centrinis cilindras.
Šaknies plaukų struktūra
Šaknų plaukeliai yra labai pailgos išorinių ląstelių, dengiančių šaknį, ataugos. Šaknų plaukelių skaičius labai didelis (nuo 200 iki 300 plaukelių 1 mm2). Jų ilgis siekia 10 mm. Plaukai susiformuoja labai greitai (jaunuose obelų daiguose per 30-40 val.). Šaknų plaukai yra trumpalaikiai. Jie žūva per 10-20 dienų, o ant jaunos šaknies dalies išauga nauji. Tai užtikrina naujų dirvožemio horizontų vystymąsi prie šaknies. Šaknis nuolat auga, formuodamas vis daugiau naujų šaknų plaukelių plotų. Plaukai gali ne tik sugerti paruoštus medžiagų tirpalus, bet ir prisidėti prie tam tikrų dirvožemio medžiagų ištirpinimo, o vėliau jas sugerti. Šaknies sritis, kurioje nunyko šaknų plaukeliai, kurį laiką gali sugerti vandenį, bet vėliau pasidengia kamščiu ir praranda savo gebėjimą.
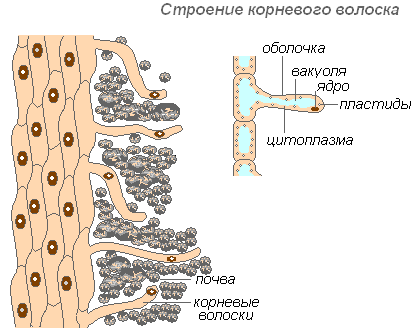
Plaukų apvalkalas yra labai plonas, o tai palengvina maistinių medžiagų įsisavinimą. Beveik visą plauko ląstelę užima vakuolė, apsupta plonu citoplazmos sluoksniu. Branduolys yra ląstelės viršuje. Aplink ląstelę susidaro gleivinis apvalkalas, kuris skatina šaknų plaukelių sulipimą su dirvožemio dalelėmis, o tai pagerina jų kontaktą ir padidina sistemos hidrofiliškumą. Įsisavinimą palengvina šaknų plaukeliais išskiriamos rūgštys (anglies, obuolių, citrinų), kurios tirpdo mineralines druskas.
Šaknų plaukeliai taip pat atlieka mechaninį vaidmenį - jie tarnauja kaip atrama šaknies viršūnei, kuri praeina tarp dirvožemio dalelių.
Po mikroskopu, šaknies skerspjūvyje absorbcijos zonoje, jos struktūra matoma ląstelėje ir audinių lygiai. Šaknies paviršiuje yra šakniastiebis, po juo – žievė. Išorinis žievės sluoksnis yra egzoderma, į vidų nuo jo yra pagrindinė parenchima. Jo plonasienės gyvos ląstelės atlieka saugojimo funkciją, veda maistinių medžiagų tirpalus radialine kryptimi – nuo sugeriančio audinio į medienos indus. Jie taip pat sintetina daugybę augalui gyvybiškai svarbių organinių medžiagų. Vidinis žievės sluoksnis yra endoderma. Maistiniai tirpalai, patenkantys iš žievės į centrinį cilindrą per endodermos ląsteles, praeina tik per ląstelių protoplastą.

Žievė supa centrinį šaknies cilindrą. Jis ribojasi su ląstelių sluoksniu, kuris ilgą laiką išlaiko gebėjimą dalytis. Tai yra periciklas. Periciklinės ląstelės sukelia šonines šaknis, priedinius pumpurus ir vidurinio ugdymo audinius. Į vidų nuo periciklo, šaknies centre, yra laidūs audiniai: bastas ir mediena. Kartu jie sudaro radialinį laidų spindulį.
Šaknies laidi sistema veda vandenį ir mineralus iš šaknies į stiebą (srovė aukštyn), o organines medžiagas iš stiebo į šaknį (srovė žemyn). Jis susideda iš kraujagyslių pluoštinių ryšulių. Pagrindiniai pluošto komponentai yra floemo (per kurį medžiagos juda į šaknį) ir ksilemo (per kurią medžiagos juda iš šaknies) atkarpos. Pagrindiniai laidūs floemo elementai yra sieto vamzdeliai, ksilemos – trachėjos (kraujagyslės) ir tracheidės.
Šaknų gyvybės procesai
Vandens transportas prie šaknų
Vandens sugėrimas šaknų plaukeliais iš dirvožemio maistinių medžiagų tirpalo ir jo laidumas radialine kryptimi išilgai pirminės žievės ląstelių per praėjimo ląsteles endodermyje į radialinio kraujagyslių pluošto ksilemą. Vandens sugėrimo šaknų plaukeliais intensyvumas vadinamas siurbimo jėga (S), jis lygus skirtumui tarp osmosinio (P) ir turgorinio (T) slėgio: S=P-T.
Kai osmosinis slėgis lygus turgoro slėgiui (P=T), tada S=0, vanduo nustoja tekėti į šaknies plauko ląstelę. Jei medžiagų koncentracija dirvožemio maistinių medžiagų tirpale yra didesnė nei ląstelės viduje, tada vanduo išeis iš ląstelių ir įvyks plazmolizė – augalai nuvys. Šis reiškinys pastebimas esant sausam dirvožemiui, taip pat naudojant per daug mineralinių trąšų. Šaknies ląstelių viduje šaknies čiulpimo galia didėja nuo šakniastiebio link centrinio cilindro, todėl vanduo juda koncentracijos gradientu (ty iš didesnės koncentracijos vietos į vietą, kurioje koncentracija mažesnė) ir sukuria šaknų slėgį. kuris pakelia vandens stulpelį išilgai ksilemo indų, sudarydamas aukštyn kylančią srovę. Jį galima rasti ant pavasarinių belapių kamienų, kai renkama „syla“, arba ant nupjautų kelmų. Vandens nutekėjimas iš medienos, šviežių kelmų, lapų vadinamas augalų „raudojimu“. Lapams žydint jie taip pat sukuria siurbimo jėgą ir pritraukia vandenį į save – kiekviename inde susidaro ištisinis vandens stulpelis – kapiliarų įtampa. Šaknies slėgis yra apatinis vandens srovės variklis, o lapų siurbimo galia yra viršutinė. Tai galite patvirtinti atlikdami paprastus eksperimentus.
Vandens sugėrimas per šaknis
Tikslas: išsiaiškinti pagrindinę šaknies funkciją.
Ką mes darom: ant šlapių pjuvenų išaugintą augalą nukratykite jo šaknų sistemą ir nuleiskite šaknis į stiklinę vandens. Užpilkite plonu sluoksniu ant vandens, kad apsaugotumėte jį nuo išgaravimo. daržovių aliejus ir atkreipkite dėmesį į lygį.

Ką mes stebime: po paros ar dviejų vandens bake nukrito žemiau žymos.

Rezultatas: todėl šaknys įsiurbė vandenį ir iškėlė jį į lapus.
Galima atlikti dar vieną eksperimentą, įrodantį maistinių medžiagų įsisavinimą iš šaknų.
Ką mes darom: nupjauname augalo stiebą, palikdami 2-3 cm aukščio kelmą.Ant kelmo uždedame 3 cm ilgio guminį vamzdelį, o viršutiniame gale uždedame lenktą stiklinį vamzdelį 20-25 cm aukščio.
Ką mes stebime: vanduo stikliniame vamzdyje pakyla ir išteka.
Rezultatas: tai įrodo, kad šaknis sugeria vandenį iš dirvos į stiebą.

Ar vandens temperatūra turi įtakos vandens įsisavinimo greičiui iš šaknų?
Tikslas: sužinoti, kaip temperatūra veikia šaknies veiklą.
Ką mes darom: viena stiklinė turi būti su šiltu vandeniu (+17-18ºС), o kita su šaltu (+1-2ºС).
Ką mes stebime: pirmuoju atveju vanduo išsiskiria gausiai, antruoju – mažai arba visiškai sustoja.
Rezultatas: tai yra įrodymas, kad temperatūra stipriai veikia šaknų veiklą.

Šiltas vanduo aktyviai įsisavinamas šaknų. Pakyla šaknų slėgis.

Šaltą vandenį prastai pasisavina šaknys. Tokiu atveju šaknų slėgis sumažėja.
mineralinė mityba
Fiziologinis mineralų vaidmuo yra labai didelis. Jie yra sintezės pagrindas organiniai junginiai, taip pat koloidų fizinę būklę keičiantys veiksniai, t.y. tiesiogiai veikia protoplasto metabolizmą ir struktūrą; veikia kaip biocheminių reakcijų katalizatoriai; paveikti ląstelės turgorą ir protoplazmos pralaidumą; yra elektrinių ir radioaktyvių reiškinių centrai augalų organizmuose.
Nustatyta, kad normalus augalų vystymasis įmanomas tik tada, kai maistinių medžiagų tirpale yra trys nemetalai – azotas, fosforas ir siera bei – ir keturi metalai – kalis, magnis, kalcis ir geležis. Kiekvienas iš šių elementų turi individualią vertę ir negali būti pakeistas kitu. Tai makroelementai, jų koncentracija augale yra 10 -2 -10%. Normaliam augalų vystymuisi reikalingi mikroelementai, kurių koncentracija ląstelėje yra 10 -5 -10 -3%. Tai boras, kobaltas, varis, cinkas, manganas, molibdenas ir tt Visų šių elementų randama dirvožemyje, tačiau kartais jų yra nepakankamai. Todėl į dirvą tręšiamos mineralinės ir organinės trąšos.
Augalas normaliai auga ir vystosi, jei šaknis supančioje aplinkoje yra visų reikalingų maisto medžiagų. Dirva yra tokia aplinka daugeliui augalų.
Šaknų kvėpavimas
Normaliam augalo augimui ir vystymuisi būtina, kad į šaknį patektų grynas oras. Patikrinkim ar taip?
Tikslas: ar šaknims reikia oro?
Ką mes darom: Paimkime du vienodus indus su vandeniu. Į kiekvieną indą dedame besivystančius sodinukus. Kiekvieną dieną viename iš indų vandenį prisotiname oru, naudodami purškimo pistoletą. Antrame inde ant vandens paviršiaus užpilkite ploną augalinio aliejaus sluoksnį, nes jis sulėtina oro patekimą į vandenį.

Ką mes stebime: po kurio laiko antrojo indo augalas nustos augti, nuvys ir galiausiai mirs.

Rezultatas: augalas miršta dėl to, kad trūksta oro, reikalingo šaknies kvėpavimui.
Šaknų modifikacijos
Kai kurių augalų atsarginės maistinės medžiagos nusėda šaknyse. Juose kaupiasi angliavandeniai, mineralinės druskos, vitaminai ir kitos medžiagos. Tokios šaknys stipriai auga ir įgauna neįprastą išvaizdą. Tiek šaknis, tiek stiebas dalyvauja formuojant šakniavaisius.
Šaknys
Jei atsarginės medžiagos kaupiasi pagrindinėje šaknyje ir pagrindinio ūglio stiebo apačioje, susidaro šakniavaisiai (morkos). Šaknis formuojantys augalai dažniausiai būna dvimečiai. Pirmaisiais gyvenimo metais jie nežydi ir sukaupia daug maisto medžiagų šakniavaisiuose. Antra, jie greitai žydi, naudodami sukauptas maistines medžiagas ir suformuoja vaisius bei sėklas.
šaknų gumbai

Jurginuose atsarginės medžiagos kaupiasi atsitiktinėse šaknyse, suformuodamos šaknų gumbus.
bakteriniai mazgeliai
Savotiškai pakitusios dobilų, lubinų, liucernos šoninės šaknys. Bakterijos nusėda jaunose šoninėse šaknyse, o tai prisideda prie dujinio azoto pasisavinimo iš dirvožemio oro. Tokios šaknys būna mazgelių pavidalo. Šių bakterijų dėka šie augalai gali gyventi azoto neturtingose dirvose ir padaryti jas derlingesnes.
dygliuotas

Potvynių zonoje auganti rampa išvysto dygliuotas šaknis. Aukštai virš vandens jie laiko didelius lapinius ūglius ant netvirtos dumblinos žemės.
Oras

Tropiniai augalai, gyvenantys ant medžių šakų, vysto oro šaknis. Jie dažnai randami orchidėjose, bromeliadose ir kai kuriuose paparčiuose. Orinės šaknys laisvai kabo ore, nepasiekdamos žemės ir sugerdamos drėgmę nuo lietaus ar ant jų krintančios rasos.
Įtraukikliai

Svogūniniuose ir svogūniniuose augaluose, pavyzdžiui, krokusuose, tarp daugybės siūlinių šaknų yra keletas storesnių, vadinamųjų besitraukiančių šaknų. Sumažėjus, tokios šaknys įtraukia gumbasvogūnius giliau į dirvą.
Stulpo formos

Fikusai vysto stulpines antžemines šaknis arba atramines šaknis.
Dirva kaip šaknų buveinė
Dirvožemis augalams yra aplinka, iš kurios jie gauna vandens ir maistinių medžiagų. Mineralinių medžiagų kiekis dirvožemyje priklauso nuo specifinių pirminės uolienos ypatybių, organizmų veiklos, pačių augalų gyvybinės veiklos, dirvožemio tipo.
Dirvožemio dalelės konkuruoja su šaknimis dėl drėgmės, išlaikydamos ją savo paviršiuje. Tai vadinamasis surištas vanduo, kuris skirstomas į higroskopinį ir plėvelinį. Jį laiko molekulinės traukos jėgos. Augalui prieinamą drėgmę atstoja kapiliarinis vanduo, kuris koncentruojasi mažose dirvožemio porose.
Antagonistiniai santykiai susidaro tarp dirvožemio drėgmės ir oro fazės. Kuo dirvoje daugiau didelių porų, tuo geresnis šių dirvožemių dujų režimas, dirva sulaiko mažiau drėgmės. Palankiausias vandens-oro režimas palaikomas struktūriniuose dirvožemiuose, kur vanduo ir oras išsidėsto vienu metu ir netrukdo vienas kitam - vanduo užpildo struktūrinių agregatų viduje esančius kapiliarus, o oras užpildo tarp jų esančias dideles poras.
Augalo ir dirvožemio sąveikos pobūdis daugiausia susijęs su dirvožemio sugeriamumu – gebėjimu išlaikyti arba surišti cheminius junginius.
Dirvožemio mikroflora daugiau skaido organines medžiagas paprastos jungtys, dalyvauja formuojant dirvožemio struktūrą. Šių procesų pobūdis priklauso nuo dirvožemio tipo, augalų liekanų cheminės sudėties, mikroorganizmų fiziologinių savybių ir kitų veiksnių. Dirvožemio struktūros formavime dalyvauja dirvožemio gyvūnai: anelidai, vabzdžių lervos ir kt.
Dėl biologinių ir cheminiai procesai dirvožemyje susidaro sudėtingas organinių medžiagų kompleksas, kurį jungia terminas „humusas“.
Vandens kultūros metodas
Kokių druskų reikia augalui ir kokią įtaką jos daro jo augimui bei vystymuisi, buvo nustatyta eksperimentuojant su vandens kultūromis. Vandens kultūros metodas – tai augalų auginimas ne dirvoje, o vandeniniame mineralinių druskų tirpale. Atsižvelgiant į eksperimento tikslą, iš tirpalo galite neįtraukti atskiros druskos, sumažinti arba padidinti jos kiekį. Nustatyta, kad azoto turinčios trąšos skatina augalų augimą, turinčios fosforo – anksčiausiai sunoksta vaisius, o turinčios kalio – greičiausią organinių medžiagų nutekėjimą iš lapų į šaknis. Šiuo atžvilgiu azoto turinčias trąšas rekomenduojama tręšti prieš sėją arba pirmoje vasaros pusėje, turinčias fosforo ir kalio – antroje vasaros pusėje.
Vandens kultūrų metodu pavyko nustatyti ne tik augalo makroelementų poreikį, bet ir išsiaiškinti įvairių mikroelementų vaidmenį.
Šiuo metu pasitaiko atvejų, kai augalai auginami hidroponikos ir aeroponikos metodais.
Hidroponika – tai augalų auginimas vazonuose, užpildytuose žvyru. Maistinis tirpalas, kurio sudėtyje yra būtini elementai, paduodamas į indus iš apačios.
Aeroponika yra augalų oro kultūra. Taikant šį metodą, šaknų sistema yra ore ir automatiškai (kelis kartus per valandą) purškiama silpnu maistinių druskų tirpalu.
ORGANOGRAFIJA
organografija - gyvo organizmo organų aprašymas.
Vargonai– morfologiškai diferencijuota ir funkciškai specializuota kūno dalis. Pagal dvi pagrindines funkcijas: palaikyti tam tikro augalo gyvybę ir daugintis keliose vėlesnėse kartose, išskiriami vegetatyviniai ir generatyviniai (dauginimosi) organai.
Pirmiausia ištirsime vegetatyvinius organus, kurie sudaro ūglių sistemą ir šaknų sistemą. Šaknų sistemos elementas yra šaknis.
Šaknis
Šaknis išsivystė vėliau nei ūglis ir kilęs iš pirmųjų sausumos augalų – rinofitų – rizoidų.
Skirtingos šaknies dalys atlieka skirtingas funkcijas ir joms būdingos tam tikros morfologinės savybės. Šios dalys vadinamos zonomis. Jaunos šaknies zonos: dalijimosi zona, augimo arba tempimo zona, absorbcijos arba absorbcijos zona ir laidumo zona.
Išorėje viršūninę meristemą dengia šaknies dangtelis. Gyvos dangtelio parenchiminės ląstelės gamina gleives. Gleivinė medžiaga yra labai hidratuotas pektino polisacharidas. Jis kaupiasi diktiozomų pūslelėse, kurios, susijungusios su plazmine membrana, išleidžia ją į tarpą tarp plazminės membranos ir ląstelės membranos. Tada gleivės patenka į ląstelės membranos paviršių, kur susidaro maži lašeliai.
Dėl šios gleivinės dangos dirvožemio dalelės prilimpa prie šaknų galiuko ir šaknų plaukelių. Gleivės tarnauja kaip sugeriantis paviršius jonų mainams. Gleivinė apsaugo šaknies galiuką nuo kenksmingų medžiagų dirvožemis ir išdžiūvimas, sukuria palankias sąlygas naudingosioms bakterijoms apsigyventi.
Kai šaknis auga ir stumia šaknies dangtelį į priekį, ląstelės, esančios kepurėlės pakraštyje, yra išardomos. Ląstelių gyvenimo trukmė nuo atsiradimo iki nuplikimo yra 4-9 dienos, priklausomai nuo kepurėlės ilgio ir augalo rūšies.
Šaknies dangtelis valdo šaknies georecepciją. Gravitacijos suvokimo vieta dangtelyje yra centrinė ląstelių kolonėlė – kolumelės, kuriose amiloplastai veikia kaip statolitai (gravitacijos receptoriai).
Viršūninę meristemą sudaro mažos (10–20 µm skersmens) daugiakampės ląstelės su tankia citoplazma ir dideliais branduoliais.
Sporiniai augalai turi monoinicialinį šaknies viršūnės tipą.
Sėklinių augalų šaknyse aprašyti du pagrindiniai viršūninio organizavimo tipai. Viename iš jų visos sritys turi bendrus inicialus. Tai atviras viršūninės meristemos tipas, kuris tikriausiai yra filogenetiškai primityvus. Antrajame tipe kiekviena iš trijų dalių, t.y. centrinis (arba laidus) cilindras, pirminė žievė ir šaknies dangtelis turi savo inicialus. Tai uždaro tipo viršūninė meristema.
Inicialai, atsakingi už ląstelių augimo organizavimą pirmaisiais etapais, paprastai praranda mitozinį aktyvumą vėlesniuose šaknų augimo etapuose. Vietoj to, ląstelės, esančios šiek tiek giliau šaknies kūne, demonstruoja mitozinį aktyvumą. Dėl šių stebėjimų susiformavo ramybės centro viršūninėje meristemoje teorija. Remiantis šia teorija, labiausiai nutolusios ląstelės (tolimiausia organo dalis, nutolusi nuo centrinio taško: apatinis šaknies galas) šaknies kūne (pirmieji pleromos ir periblemos inicialai) dalijasi retai, turi maždaug vienodas. dydis ir mažas NA bei baltymų sintezės greitis. Poilsio centras yra pusrutulio arba disko formos, skiriasi tūriu, priklausomai nuo šaknies dydžio. Poilsio centro ląstelės dalijasi maždaug 10 kartų lėčiau nei kaimyninės ląstelės.
Eksperimentiškai pažeistose šaknyse ramybės centras gali atkurti meristemą. Tai yra, poilsio centras vaidina esminį vaidmenį formuojant struktūrą ir plėtojant šaknį.
Šaknies galiuko ypatybė yra išilginių ląstelių eilių, kilusių iš viršūninės meristemos, išdėstymas.
Šaknies viršūninės meristemos sluoksniai:
Dermatogenas(gr. derma – oda, genos – gimimas) – išorinis viršūninės meristemos sluoksnis, diferencijuojasi į rizodermą. Tinkamos formos dermatogeninės ląstelės.
Pleroma(gr. pleroma – užpildymas) – centrinė viršūninės meristemos dalis, diferencijuojasi į centrinio cilindro ląsteles. Ląstelės yra pailgos išilgai ašies.
periblem(gr. periblema – danga) – viršūninės meristemos sluoksnis tarp dermatogeno ir pleromos, diferencijuojasi į pirminės žievės ląsteles.
Pirminė žievė diferencijuojasi pailgėjimo zonos lygyje. Kolonizacijos zonoje pirminė žievė sudaro didžiąją skerspjūvio dalį. Jis vadinamas „pirmine“ dėl kilimo iš pirminės meristemos, „žieve“ – dėl periferinės padėties.
Pirminės žievės išorinės ląstelės, esančios po rizoderma, vadinamos egzoderma. Pirminės žievės vidinis sluoksnis yra endoderma.
Didžioji pirminės žievės dalis yra mezoderma. Mezodermą sudaro laisvai išsidėsčiusios parenchiminės ląstelės, kurios prisideda prie šaknies ląstelių aeracijos.
Gimnosėklių ir daugumos dviskilčių šaknys anksti numeta pirminę žievę. Juose pirminės žievės ląstelės išlieka parenchiminės. Priešingai, pirminė vienaląsčių žievė išlieka per visą šaknies gyvenimą, o jos ląstelės sudaro antrinius lignifikuotus apvalkalus.
Šaknies centrą užima ašinis cilindras – stela. Pradeda diferencijuotis augimo zonoje. Ašinio cilindro formavimasis prasideda nuo jo išorinio sluoksnio susidarymo - periciklas. Periciklas priklauso pirminėms šoninėms meristemoms, jo ląstelės ilgą laiką išlaiko meristeminį aktyvumą. Paprastai periciklą vaizduoja vienas ląstelių sluoksnis, nors jis gali būti ir daugiasluoksnis.
Daugumoje sėklinių augalų šoninės šaknys klojasi pericikle, atsiranda priedinių pumpurų užuomazgos. Antrinio augimo rūšyse periciklas dalyvauja formuojant kambį ir sukuria pirmąjį felogeno sluoksnį.
Po periciklu dedamos prokambio ląstelės, iš kurių susidaro pirminė floema, vėliau – pirminė ksilema. Įvairiose rūšyse ksilemo spindulių skaičius skiriasi nuo dviejų, trijų, keturių iki kelių. Pagal ksilemo spindulių skaičių šaknys vadinamos diarchu, triarchu, tetraarchu, poliarchu. Tetrarcho šaknys laikomos pagrindinėmis. Vienakojai turi poliarchinę stelą.
Šiuo būdu, bendras planas Pirminė šaknies struktūra ir morfologinė audinių diferenciacija rodo aukštą šaknies specializacijos lygį ir prisitaikymą prie vandens ir mineralų absorbcijos ir laidumo funkcijų.
Šoninių šaknų formavimasis
Šoninės šaknys atsiranda centrinio cilindro periferijoje skirtingais atstumais nuo viršūninės meristemos. Nes jie dedami į giliai esančius šaknies audinius, jų kilmė vadinama endogenine (iš graikų kalbos „gimęs viduje“). Periciklėje klojamos šoninės gimnasėklių ir gaubtasėklių šaknys. Šoninių šaknų formavime dalyvaujančios periciklo ląstelės yra šaknies lankas.
Endoderma taip pat gali sudaryti kelis šaknies primordijų ląstelių sluoksnius.
Diarchinėje šaknyje tarp floemos ir ksilemo klojama šoninė šaknis, triarchinėje, tetrachninėje šaknyje priešais ksilemą, poliarchinėje vienakilčių šaknyse – priešais floemą. Periciklinės ląstelės dalijasi įvairiomis kryptimis. Dėl to susidaro mažas šaknies gumbas. Didėjant dydžiui, šaknies rudimentas spaudžia endodermą. Tačiau endodermos apvalkalai yra kieti. Todėl endoderma išsikiša. Endodermos iškilimas vadinamas šaknies kišene.
Šaknies pradas praeina per pirminę žievę, išskirdamas fermentus, kurie pakeliui naikina žievės ląsteles. Ankstyvosiose vystymosi stadijose šis užuomazgas sudaro šaknies kepurėlę, viršūnes ir pirmines meristemas. Laikui bėgant, dėl parenchiminių ląstelių diferenciacijos į ksilemo ir floemo elementus, centriniai šoninių ir pirminių šaknų cilindrai yra tarpusavyje susiję.
Vienakilčių pirminė šaknų struktūra išsaugoma visą gyvenimą ir antriniai audiniai neatsiranda. Su amžiumi gali padidėti ląstelių dydžiai, storėti ląstelių membranos, tačiau bendras šaknies struktūros „planas“ išlieka nepakitęs.
Gimnosėkliuose ir dviskilčiuose atsiranda antrinių pakitimų, susidaro antrinė šaknų struktūra. Šaknies antrinės struktūros formavimasis pirmiausia yra susijęs su kambio veikla. Kambis susidaro dalijantis toms prokambio ląstelėms, kurios lieka nediferencijuotos tarp pirminės floemos ir pirminės ksilemos. Tada periciklo ląstelės, esančios už ksilemo iškyšų, taip pat demonstruoja kambinį aktyvumą.
Kambis nusodina antrines ksilemo ląsteles į centrą, o antrines floemo ląsteles - į periferiją. Taip susidaro laidūs ryšuliai. Jų susidaro tiek, kiek buvo pirminės ksilemos spindulių. Esant augančių antrinių audinių spaudimui, pirminės floemos ląstelės išsilygina. Pirminis ksilemas išlaiko savo centrinę padėtį ir yra matomas kaip žvaigždutė. Laikui bėgant, pirminis ksilemas taip pat išnyksta.
Antrinėje šaknies struktūroje pirminiai audiniai nefunkcionuoja kaip laidūs elementai. Medžiagų srovė abiem kryptimis vykdoma per antrinius audinius.
Kambis, kilęs iš periciklo, sudaro plačius parenchiminius spindulius, esančius tarp atvirų kolateralinių pluoštų. Tai yra pirminiai šerdies spinduliai. Jie užtikrina fiziologinį ryšį tarp centrinės šaknies dalies ir pirminės žievės. Vėliau gali susidaryti siauresni antriniai meduliniai spinduliai, jungiantys antrinį ksilemą ir floemą.
Antriniai pokyčiai vyksta ne tik centriniame cilindre. Padidėjus laidžių audinių storiui, pirminė žievė išstumiama į išorę, plyšta ir išsilieja kartu su endoderma.
Išorinėje periciklo dalyje felogenas kyla ir nusėda į išorę, o daugiasluoksnę - į vidų. Taigi dviskilčių šaknys, turinčios antrinę struktūrą, yra padengtos periderma.
Daugiametėse šaknyse felogenas atsiranda giliai šaknyje. Tada šaknis, kaip ir stiebas, suformuoja ritmiką.
Svarbus antrinio šaknų augimo ekologinis aspektas yra skirtingų tos pačios rūšies augalų šaknų gebėjimas augti natūraliai. Ten, kur šaknys liečiasi viena su kita, jos susijungia per antrinį augimą. Kai šaknys auga kartu, nustatomas laidžiosios sistemos tęstinumas tarp abiejų susiliejusių partnerių. Dėl šaknų susiliejimo daugelis medžių yra tarpusavyje sujungti, o jei kai kurie iš jų nukertami, kelmai išlieka gyvi ilgą laiką. Šaknų susiliejimas prisideda prie greito infekcijos perdavimo iš vieno medžio į kitą.
Šaknų diferenciacija šaknų sistemose pasireiškia įvairiomis formomis. Didžiausią reikšmę turi diferencijavimas į pagrindines, šonines ir atsitiktines šaknis.
Specialioje grupėje išskiriamos šaknys, kylančios aukštesniųjų sporinių augalų viršūninėse meristemose. Daroma prielaida, kad jos yra archajiškiausia šaknų grupė ir išlaikė didžiausią panašumą su pirminėmis šaknimis, kurios buvo vienos išsišakojusios antžeminių ir požeminių ašių sistemos dalis. Į šią grupę įtrauktos šaknys vadinamos apikogeninėmis.
Remdamiesi šaknų sistemų atsiradimo seka evoliucijos procese ir priklausymu didelėms sisteminėms grupėms, galime apibūdinti tris evoliucinis šaknų sistemų lygis.
I. Aukštesnių sporų šaknų sistemas atstovauja tik apigeninės šaknų sistemos, paeiliui pakeičiančios viena kitą, o tai yra pagrindinis šaknų sistemų atsinaujinimo mechanizmas. Išsišakojimas yra viršūninis arba šoninis, ir gali būti, kad abu šakų tipai atsirado nepriklausomai vienas nuo kito. Nėra metamorfozių ir šaknų su siaura specializacija.
P. Gimnosėklių šaknų sistema išsiskiria pagrindinės šaknies, kuri gali būti siejama su sėklos atsiradimu, ir dvipolio embriono su embrionine šaknimi buvimu. Į atsinaujinimo mechanizmą buvo įtrauktos ne tik pavaldžių kategorijų šoninės šaknys, bet ir rizogeninės papildomos šaknys (t. y. atsitiktinės šaknys ant šaknų, papildomos šaknys ant ūglių gimnasėkliuose nebuvo pastebimai išsivystę). Specializuotos ir metamorfinės šaknys yra retos.
Sh.Angiasėklių šaknų sistemai būdingas stiprus atsitiktinių šaknų išsivystymas, metamorfozės ir siauros specializacijos šaknys. Atnaujinimo mechanizmai yra įvairūs ir veiksmingi.
Šaknų sistemų metamorfozės
Šaknų sistemų metamorfozės yra anatominių ir morfologinių ypatybių rinkinys, išsivystęs filogenetiškai ir paveldimai, fiksuotų ir sukeltų fiziologinių funkcijų pasikeitimo. Metamorfozė (metamorfozė, graikiškai – transformacija) yra genetiniame lygmenyje įvykusių pokyčių, lemiančių organo, organų sistemos ar viso organizmo morfologinį ir funkcinį pertvarkymą, rezultatas.
I. Metamorfozės, susijusios su netipinių funkcijų atlikimu:
1) kvėpavimo šaknys;
2) epifitų oro šaknys;
3) dygliuotos šaknys;
4) atsarginės šaknys.
P. Metamorfozės, susijusios su hiperfunkcija:
1) šakniavaisiai;
2) šaknų spurgai;
3) šaknies mazgeliai;
4) mikorizė.
Šakniavaisiai būdingi dvimečiams dviskilčiams augalams. Tai laikinas pavadinimas. Jie išsivysto iš vegetatyvinių organų ir neturi nieko bendra su vaisiais. Apatinė stiebo dalis ir šaknis dalyvauja formuojant šakniavaisį.
Pirmaisiais gyvenimo metais augalas vegetuoja, formuojasi šakniavaisiai. Antraisiais metais formuojasi žiedai, vaisiai, sėklos. Tai reiškia, kad šakniavaisiai yra medžiagų tiekimo organas, leidžiantis augalui perkelti ramybės periodą ir visiškai vystytis kitiems metams.
Šaknų tipai:
1.Monomambialinis.
Pradinė šaknis yra ašinis cilindras. Šakniavaisių formavimasis prasideda nuo antrinių pokyčių, susijusių su vieno kambarinio žiedo atsiradimu ir veikimu bei tolimesne parenchimatizacija.
Žinome, kad kambis antrinę ksilemą deda į vidų, o antrinę floemą – į išorę. Anatominiai skirtumai yra susiję su saugojimo audinio vystymosi vieta. Jei floemoje atsiranda stiprus parenchimos vystymasis, susidaro floeminis šakniavaisių tipas – morkų tipas. Jei ksileme - ksilemo tipas - ropės tipas.
2.Polikambinis šakniavaisių tipas.
Susidaro vienas tipinis kambalinis žiedas, likusieji – papildomi periciklinės kilmės. Jų gali būti iki 25. Šie papildomi kambio žiedai sudaro atvirus kolateralinius kraujagyslių pluoštus. Kuo toliau žiedas yra nuo pirmojo kambarinio žiedo, tuo daugiau saugojimo parenchimo jis sukuria. Daugiakampių šakniavaisių pavyzdys yra burokėliai.
Vienkambariuose ir daugiakambariuose šakniavaisiuose pirminė žievė anksti žūva ir pleiskanoja. Dengiantis audinys yra periderma.
Taigi bendras visų šakniavaisių bruožas yra didelis saugojimo parenchimos kiekis, persmelktas laidžių audinių. Glaudus ryšys tarp laidžių ir saugojimo audinių pasiekiamas įvairiais antrinio augimo modifikacijomis.
Kai kuriuose augaluose sandėliavimo šaknys susidaro iš šoninių ir atsitiktinių šaknų šaknų kūgių pavidalu. Vienaląsčių (lyubkos) šaknų kūgiai yra kambaliniai, turi pirminę struktūrą; dviskilčiai – kambaliniai, turi antrinę struktūrą. Pavyzdys: floem tipo jurginų šaknų spurgai.
Šaknų spurgai sudaro atsitiktines šaknis visame paviršiuje, o viršutinėje dalyje - atsitiktinius pumpurus, todėl yra vegetatyvinio dauginimosi organai.
Žinoma, kad augalų augimą dažniausiai riboja nepakankamas fiksuoto azoto tiekimas. Azotas, esantis stabilios molekulės N 2 pavidalu, sudaro 80% atmosferos. Prieš fiksavimą (N 2 redukavimą iki amoniako NH 3) ši molekulė turi būti suskaidyta. Azoto fiksavimą atlieka kai kurios laisvai gyvenančios bakterijos, kurios sunaudoja dirvožemio organines medžiagas. Kai kurios bakterijos nusėda ant aukštesnių augalų šaknų. Organinėmis šaknų išskyromis augalas „pritraukia“ bakterijas. O mikroorganizmai savo ruožtu aprūpina augalą fiksuotu azotu. Ši abipusiai naudinga dviejų organizmų asociacija vadinama simbioze. Dažniausiai šaknys jungiasi su azotą fiksuojančiomis Rhizobium genties bakterijomis. Daugiau nei 60 dviskilčių ir vienaskilčių augalų šeimų gali užmegzti simbiozę su bakterijomis. Kiekviena augalų rūšis turi savo simbiotinį Rhizobium. Abipusis augalo ir bakterijos atpažinimas atliekamas prie konkrečios bakterijos prijungiant specialų baltymą – lektiną, esantį trichoblastų paviršiuje. Šaknų plaukeliai, veikiami bakterijų išskiriamų augimo hormonų, deformuojasi. Bakterijos patenka į šias susuktas ląsteles. Ląstelių viduje bakterijos dauginasi, sudarydamos infekcinį siūlą, uždarytą dantenų apvalkale. Šis siūlas įsiskverbia giliai į šaknį. Tokio bakterijų įsiskverbimo rezultatas yra stiprus šaknų mezodermos ląstelių dauginimasis, dėl kurio susidaro mazgeliai. Tolimiausia mazgo dalis ilgą laiką išlieka meristemine zona, o vidinėje mazgo dalyje yra bakterijos. Pats bakterinis audinys yra mažos ląstelės, turinčios klampią citoplazmą ir daugybę bakterijų. Bakterinis audinys yra apsuptas kraujagyslių pluoštų, kurie yra sujungti su centriniu šaknies cilindru. Tie. yra transporto sistema, skirta maisto medžiagų mainams tarp bakterijų ir augalų. Azotą fiksuojančios bakterijos teigiamai veikia augalų mitybą azotu ir dirvožemio derlingumą.
Fiziologinį šaknies aktyvumą, susijusį su augalo aprūpinimu vandeniu ir mineralais, gali sustiprinti jos simbiotinis ryšys su konkrečiu grybu. Tokia glaudi abipusiai naudinga šaknų ir grybų simbiozė yra mikorizė. Mikorizė yra žinoma daugumoje kraujagyslių augalų. Tik keletas žydinčių šeimų jo nesudaro arba formuojasi retai, pavyzdžiui, kopūstai, viksvos.
Mikoriziniai grybai atlieka pagrindinį vaidmenį kraujagyslių augalų mineralinėje mityboje. Reikšmingas mikorizės vaidmuo absorbuojant ir pernešant fosforą.
Dvišalio transportavimo metu (fosfatų perkėlimas iš grybų į augalus ir angliavandeniai priešinga kryptimi) apoplastinėje zonoje tarp augalo ir grybelio plazminių membranų atsiranda nuostolių ir absorbcijos.
Mikoriziniai grybai naudoja P šaltinius, kurie paprastai nėra prieinami augalams. Mikorizė taip pat prisideda prie augalų Zn, Mn, Cu pasisavinimo. Šie elementai dirvožemyje yra silpnai judrūs, todėl aplink šaknų plaukelius greitai susidaro jų išeikvotos zonos. Mikorizinių grybų hifų tinklas tęsiasi kelis centimetrus nuo kiekvienos šaknies ir taip padidina šaknų sugeriamąjį paviršių.
Yra šie mikorizės tipai:
1) ektomikoriza: grybelis storu dangalu padengia visą šaknies galiuką ir prasiskverbia į tarpląstelinius tarpus;
2) endomikorizė: grybelis įsiveržia į ląsteles.
Ektomikorizė būdinga tam tikroms medžių ir krūmų šeimoms (pušims, gluosniams, bukams). Ektomikorizė susidaro ant jaunų, dar nesubyrėjusių šaknų. Ektomikorizės šaknys yra trumpos, išsišakojusios ir atrodo patinusios.
Tokiose šaknyse nuslopinamas šaknų plaukelių vystymasis, gali sumažėti viršūninės meristemos ir šaknies kepurėlės tūris. Šaknų plaukelių funkciją atlieka hifai.
Endomikorizinės šaknys savo forma panašios į paprastas šaknis, tačiau skiriasi nuo jų tamsesne spalva. Grybeliniai hifai prasiskverbia į šaknų žievės ląsteles, kur suformuoja pūsleles (pabrinkimus) ir arbuskules (į medį panašias šakas). Todėl endomikoriza dažnai vadinama vezikuline-arbuskuline. Tam tikromis sąlygomis grybelio hifus gali „suvirškinti“ ląstelės. Išsaugomi šaknų plaukai ant šaknų. Tai yra labiausiai paplitusi mikorizės rūšis. Aptinkama maždaug 80 % visų kraujagyslių augalų, būtent žoliniuose augaluose, įskaitant javus; prie krūmų, medžių (obelių, klevų...). Pavyzdžiui, jauname tuopos augale susiformuoja vezikulinė-arbuskulinė endomikoriza, o jai senstant išsivysto apvalkalo ektomikorizė.
Viržiams Ericaceae būdinga erikoidinė-arbutoidinė mikorizė. Grybelis aplink šaknį sudaro galingą hifų sankaupą, o ploni šoniniai hifai prasiskverbia į šaknies žievės ląsteles. Šie mikoriziniai grybai labai sumažina dirvožemio toksiškumą viržiams, todėl šie augalai gali įsikurti rūgščioje, maistinių medžiagų neturtingoje dirvoje.
Yra ir kita rūšis – orchidėjų endomikorizė. Orchidėjų sėklos gamtoje sudygsta tik esant atitinkamiems grybams.
Apskritai, natūraliomis sąlygomis mikorizės asociacijos yra taisyklė, o ne išimtis.
Seniausių kraujagyslinių augalų liekanų tyrimas parodė, kad endomikorizė juose pasitaikė ne rečiau nei šiuolaikiniuose augaluose. Tai leido daryti prielaidą, kad mikorizės atsiradimas evoliucijos procese gali būti vienas iš veiksnių, leidusių augalams kolonizuoti žemę. Atsižvelgiant į silpną dirvožemio vystymąsi šios kolonizacijos metu, galima daryti prielaidą, kad mikoriziniai grybai buvo augalų mineralinės mitybos pagrindas. Šiuolaikiniai pionieriniai augalai, sergantys mikorize, turi didesnes galimybes išgyventi skurdžiose dirvose.
Augalai, turintys mikotrofinį mitybos tipą, yra atsparesni sunkiųjų metalų buvimui technogeninėje aplinkoje.
Keletas žodžių apie atogrąžų miškų augalų šaknis. Dažniau nei dygliuotos, lentą primenančios atraminės šaknys. Šaknys primena lentas, kurios viršutiniais galais remiasi į medį. Nors dygliuotos šaknys yra atsitiktinės šaknys, atsirandančios ant kamieno ir augančios žemyn, lentų šaknų formavimasis yra susijęs su šoninėmis šaknimis, augančiomis tiesiai po dirvožemio paviršiumi. Šoninės šaknys vysto vertikalias, trikampio skerspjūvio ataugas, kurios yra greta medžio kamieno.
Lentų šaknų mediena dažnai būna kietesnė nei likusi to paties medžio mediena, dažnai jos būna padengtos plonu žievės sluoksniu. Sprendžiant iš šių šaknų skerspjūvyje matomų augimo žiedų, iš pradžių tai yra normalios, suapvalėjusios šaknys, tačiau vėliau susidaro labai stiprus sustorėjimas viršutinėje pusėje. Lentų šaknų skaičius ant medžio svyruoja nuo 1 iki 10, tačiau dažniausiai būna trys ar daugiau. Tropikuose lentinių šaknų formavimasis ypač pastebimas pelkėtuose miškuose ir prastai nusausintose dirvose augančiuose medžiuose. Dažnai lentos formos šaknys naudojamos pramoniniais tikslais. Taigi, Indonezijoje augančio Koompassia excelsa medžio šaknys, panašios į lentas, naudojamos valgomojo stalams gaminti.
PABĖGIMAS
Ūglis, kaip ir šaknis, yra pagrindinis augalo vegetatyvinis organas. Ūglis atlieka oro mitybos funkciją, gali atlikti daugybę kitų funkcijų ir gali metamorfozei.
Vegetatyvinis nemodifikuotas ūglis susideda iš ašinės dalies – stiebo; lapai ir pumpurai. Pagrindinis išorinis požymis, skiriantis ūglį nuo šaknies, yra ūglio lapiškumas.
Stiebo dalis, susijusi su šoninių organų (šakų, lapų ir kt.) atsiradimu - mazgas. Stiebo atkarpa tarp gretimų mazgų tarpubambliai. Vidinis kampas tarp lapo ir stiebo lapų pažastis. Kiekvienas pasikartojantis ūglio elementas (mazgas su lapu ir pumpuru lapo pažastyje ir tarpmazgas) susiformuoja metameras. Taigi šūvis susideda iš eilės metamerų, t.y. turi metamerinę struktūrą.
XX amžiaus 90-ųjų pradžioje vietoj mazgo ir tarpmazgo sąvokų amerikiečių mokslininkai pasiūlė sąvoką - stiebo elementas. Stiebo elementas yra stiebo atkarpa tarp lapo pradmenų kraštų. Atsiskleidė ūglio viršūnėje, kur dar nėra tarpubamblio. Tie. nauja „kamieno elemento“ sąvoka atitinka „metamero“ sąvoką.
Ūglio viršūninės meristemos struktūra yra sudėtingesnė nei šaknies. Iš tiesų, ūglyje viršūninė meristema ne tik formuoja pirminių audinių ląsteles, bet ir dalyvauja formuojant lapų pradmenis ir pažastinius pumpurus, kurie vėliau išsivysto į šoninius ūglius. Šios meristemos neapsaugo tokia struktūra kaip šaknies dangtelis.
Viršūnių tipai yra tokie patys kaip ir šaknyje: monoinicialus, zoninis, pakopinis. Pakopų skaičius yra didesnis nei šaknyje.
Evoliuciškai primityvi monoinitial viršūnė. Šio tipo viršūnių pavyzdys yra asiūklių. Viena didelė pailga viršūninė ląstelė yra stipriai vakuolizuota. Jis dalijasi kryptimis, lygiagrečiomis jo paviršiams (išskyrus atokiausią paviršių). Artimiausios gautos ląstelės taip pat yra labai vakuoliuotos, tačiau joms dalijantis atsiranda mažos ląstelės su tankiais protoplastais. Tokios ląstelės randamos palei stiebo viršūnės periferiją, kur dedami lapų pradmenys.
Daugumos žydinčių augalų ūglio vegetatyvinė viršūnė yra pastatyta kaip tunika - kūnas - dvi zonos, kurios skiriasi ląstelių dalijimosi plokštumomis. Tunika(lot. – apvalkalas) apima išorinius ląstelių sluoksnius, kurie dalijasi antikliniškai (statmenai paviršiui) ir suteikia paviršiaus padidėjimą. Rėmas(lot. - kūnas) - centrinė ląstelių grupė po tunika, dalijasi skirtingose plokštumose, sudarydama pagrindinį šūvio tūrį. Tunikos korpusas ir kiekvienas sluoksnis turi savo inicialus. Tunikoje inicialai dedami centre išilgai ašies. Dvikilčiai daugiausia turi dvisluoksnę, vienaląsčiai – vienasluoksnę, rečiau – dvisluoksnę.
Nors tunikos korpuso koncepcija padeda suprasti ląstelių vietą ir augimą pačioje viršūninėje meristemoje, skirtingų ląstelių zonų gretimose srityse atpažinimas padeda atskleisti meristemos indėlį į audinių ir organų diferenciaciją. šaudymas. Viršūninėje meristemoje yra skirtingų citologinių savybių turinčių zonų citohistologinis zonavimas.
Korpusas atitinka ląstelių sritį, vadinamą centrinių motininių ląstelių zona.Šios zonos ląstelės yra labai vakuoliuotos, o tai siejama su santykinai mažu mitozinio aktyvumo lygiu. Centrinių motininių ląstelių zona yra apsupta periferinės meristemos, kuri iš dalies kyla iš tunikos, iš dalies iš korpuso. Protodermos histogenas kilęs iš išorinio tunikos sluoksnio. Periferinė meristema kyla iš šoninių viršūninių inicialų darinių ir iš centrinių motininių ląstelių. Pagrindinė meristema susidaro dėl dalijimosi išilgai centrinių motininių ląstelių periferijos sluoksnyje, vadinamame pereinamoji zona. Periferinės zonos ląstelės turi tankius protoplastus, todėl turi didelį mitozinį aktyvumą.
Tam tikrose ypač aktyviose vietose dedami lapų gumbai (arba pirminiai gumbai).
Naujų lapų primordijų atsiradimas siejamas su ląstelių dalijimosi krypties pasikeitimu, prieš kurį pakinta mikrotubulių orientacija periferiniuose citoplazmos sluoksniuose. Osmosinis slėgis taip pat turi įtakos pirmtakų augimui.
Aktyvaus augimo laikotarpiu ūglio viršūninė meristema taip greitai suformuoja lapų pradmenis, kad iš pradžių mazgai ir tarpubambliai nesiskiria. Palaipsniui sritys tarp lapų pakopų pradeda augti, išsitiesti ir įgauti tarpubamblių formą. Tuo pačiu metu lapų tvirtinimo vietos formuojasi į mazgus.
Žemiau, kur atsiranda lapų užuomazgos, stebima vakuolizacija. Tai rodo pirminės žievės ir šerdies diferenciacijos pradžią. Stipri vakuolizacija pirminėje žievėje ir šerdyje rodo, kad yra meristeminė zona, vadinama likutine meristema. Atrodo, lyg tai būtų viršūninės meristemos liekana, išsilaikiusi tarp labiau diferencijuotų audinių. Žemiau lapo rudimento padėjimo vietos liekaninėje meristemoje, išilginiais padalijimais, be vėlesnio ląstelių augimo į plotį, susidaro kiek pailgos pirmojo prokambiumo ląstelės.
Kituose vystymosi etapuose liekamosios meristemos išsiskiria vis daugiau procambialinių gijų. Po to, kai susiformuoja visi tam tikro lygio stiebo kraujagyslių ryšuliai, likutinė meristema diferencijuojasi į tarpfaskulinę parenchimą. Mazguose dalis liekamosios meristemos tampa lapo plyšio parenchima.
Prokambiumas gali būti klojamas ištisiniu žiedu arba atskiromis sruogomis. Jei prokambiumas yra ištisiniame žiede, a besijos (ne ryšulio) tipo stiebo sandara. Jei prokambiumas klojamas atskiromis gijomis, a sijos tipo stiebo struktūra. Todėl pirminiai laidūs audiniai stiebe išsidėstę įvairiai: pirmuoju atveju – ištisiniame žiede, antruoju – atskiruose laidžiuose ryšuliuose, atskirtuose pagrindinės parenchimos atkarpomis – šerdies spinduliais.
Šalutiniuose kraujagyslių ryšuliuose pirminė floema atsiranda išorinėje prokambarinio laido dalyje, o pirminė ksilema – vidinėje. Tolesnė floemos diferenciacija vyksta įcentriškai: nauji floemos elementai atsiranda arčiau stiebo centro. Ksilemas diferencijuojasi priešinga kryptimi – išcentriškai.
Visa stela arba centrinė stiebo dalis susideda iš laidžių audinių, šerdies (kartais suyra) ir periciklo. Už periciklo yra pirminė žievė. Pirminės žievės (ir ji susidaro iš periferinės meristemos) sudėtis apima chlorenchimą, kartais sekrecinius elementus. Tolimiausias periferinės meristemos sluoksnis nesudaro egzodermos, kaip šaknyje, o diferencijuojasi į kolenchimą, kuris yra arba sruogų pavidalu, arba ištisiniame žiede (priklausomai nuo stiebo struktūros tipo: ryšulių ar be ryšulių). Vidinis pirminės žievės sluoksnis yra endoderma. Tačiau stiebe jis neatlieka reguliavimo funkcijos, kaip šaknyje. Krakmolo grūdeliai nusėda endodermoje, todėl jis vadinamas krakmolingu apvalkalu. Kartais jame yra kalcio oksalato kristalų.
Periciklas greitai praranda meristeminį aktyvumą ir virsta sklerenchima, kuri susidaro ištisiniu sluoksniu arba sruogomis, priklausomai nuo stiebo sandaros tipo.
Šerdį sudaro plonasienės parenchiminės ląstelės, kuriose kaupiasi atsarginės maistinės medžiagos. Yra idioblastų ląstelių, užpildytų kristalais, gleivėmis ir kt. Periferinė šerdies dalis vadinama perimeduliarinė zona(graikiškai peri – aplink, medulla – šerdis). Perimedulinės zonos ląstelės yra mažesnės ir ilgą laiką išlaiko gyvybinę veiklą.
Vienakilčiuose pradinis tipas yra stiebo struktūros ryšulio tipas, t.y. prokambiumas glūdi sruogose. Tačiau antrinė meristema – kambis – niekada neatsiranda, todėl antrinių laidių audinių nėra. Net ir sustorėjus, stiebą formuoja tik pirminiai audiniai. Vienaskilčių (delnų, alavijų) storas stiebas išsivysto dėl pagrindinės parenchimos ląstelių dalijimosi ir ištempimo. Šis augimas vadinamas difuziniu antriniu augimu.
Vienaląsčiams pirminė žievė ir centrinis cilindras nėra atskirti aiškia riba, kaip dviskilčių, o stela kyla po epidermiu.
Vienaląsčiams būdinga difuzinio tipo stiebo sandara, kai po visą stiebą išsibarstę uždari kraujagyslių ryšuliai. Tarpmazgių pailgėjimo procese pirminiai floemo ir ksilemo elementai (protoflemas ir protoksilemas) yra ištempiami ir sunaikinami, todėl ryšulio ksilemo pusėje susidaro didelė orą laikanti ertmė. Todėl laboratorinių tyrimų metu, kai pažvelgėme į ryšulių tipus, nuolat ruošiant kukurūzų stiebo skerspjūvį, buvo aišku, kad subrendęs kraujagyslių pluoštas apima du plačius metaksilemo indus, oro ertmę ir metafloemą. Kiekvienas pluoštas yra uždengtas sklerenchimos ląstelių pamušalu.
Vienaląsčiams stiebo šerdis dažnai žūva, o jos vietoje susidaro ertmė. Toks tuščiaviduris stiebas su aiškiai apibrėžtais mazgais vadinamas šiaudų. Solomina pasižymi stipriu mechaninio audinio išsivystymu, kuris stiebo periferijoje sudaro žiedą. Brandžiuose augaluose chlorenchima sunaikinama, o šiaudai įgauna sklerenchimai būdingą aukso geltonumo spalvą. Apskritai vienaląsčiams būdingas stiprus stiebo sklerifikacija, kai iki 30 % visų audinių yra sklerenchimos, gali būti dalinis gyvų parenchiminių ląstelių membranų suliginėjimas.
Dviskilčių stiebo antrinis augimas yra kambio veiklos rezultatas. Kambis pasirodo pirmųjų ūglių augimo metų pabaigoje ir prisideda prie stiebo sustorėjimo. Kambis iš dalies susidaro iš prokambiumo kraujagyslių ryšuliuose ir iš dalies iš tarpfaskulinės parenchimos. Priklausomai nuo susidarymo vietos, kambio atkarpos vadinamos fascikuliniu ir interfaskuliniu kambiu. Būsimos kambinės ląstelės tarpfascikulinėse zonose nustatomos dar ilgai prieš prasidedant kambariniam aktyvumui. Kambis pirmiausia susidaro ląstelėse, esančiose greta fasciculus, tada plinta giliai į meduliarinį spindulį. Dėl to tarpfaskulinis ir fascikulinis kambis sudaro ištisinį kamieninį žiedą stiebe.
Yra trys antrinio augimo tipai:
1) Aristolochia chircason tipas.
Inicialas yra stiebo konstrukcijos sijos tipas. Pirminiai kraujagyslių pluoštai yra atskirti plačiais šerdies spinduliais. Sujungtas kambis sudaro antrinius laidžius audinius, o tarpfaskulinis kambis – spinduliuojančią parenchimą. Ši sijos struktūra yra išsaugota.
2) Helianthus saulėgrąžų tipas.
Sijos tipo konstrukcijos taip pat yra pradinis. Tačiau antrinius laidžius audinius sudaro ir fascikulinis, ir tarpfaskulinis kambis. Antriniai kraujagysliniai ryšuliai, suformuoti tarpfaskulinio kambio, skiriasi nuo pirminių mažesniu dydžiu ir tuo, kad virš ryšulio nėra periciklinės kilmės mechaninio audinio. Ilgai veikiant kambiui, atsiranda vis daugiau antrinių ryšulių. Jie auga, susilieja vienas su kitu, sudarydami ištisinį antrinių laidžių audinių žiedą.
3) Tilia liepų tipas.
Pradinis tipas yra besijinis stiebo struktūros tipas. Bendrasis kambinis žiedas sudaro antrinius laidžius audinius. Tačiau laikas nuo laiko kambis sudaro parenchimines ląsteles, kurios toliau diferencijuojasi į antrinius spindulius.
stiebo struktūra daugiamečiai augalai
Sumedėjusiuose ir krūminiuose dviskilčiuose, taip pat spygliuočiuose antrinis tankėjimas tęsiasi daugelį metų, kai kurių rūšių kamienai siekia kelių metrų skersmenį. Sustorėjimai siejami su ilgai veikiančio kambio ir iš dalies felogeno – dviejų antrinių šoninių meristemų – veikla. Topografiškai daugiamečio sumedėjusio augalo stiebe galima išskirti tris pagrindines dalis: žievę, medieną ir šerdį. Riba tarp žievės ir medienos eina išilgai kambio.
Kambio ląstelės, skirtingai nei įprastos meristeminės ląstelės, visada yra labai vakuolizuotos. Kambinių ląstelių citoplazmos būklė kinta priklausomai nuo metų laikų.
Kambialinėje zonoje arba kambialiniame žiede aptinkamos dvi kambinių ląstelių formos: verpstės formos ląstelės, pailgintos vertikalia kryptimi (inicialai) ir sferinės ląstelės, pailgos horizontalia kryptimi – spindulių inicialai. Jei veleno ląstelės yra išdėstytos taisyklingomis horizontaliomis eilėmis, kambis vadinamas pakopiniu kambiu. Jei tokios tvarkos verpstės formos langelių išdėstyme nėra, kambis yra nepakopinis.
Iš ūdų kambio susidaro mediena ir antrinė ūda. Nepakopinis kambis sukuria neaukštą medieną ir neaukštą antrinę floemą. Evoliuciniu požiūriu ūdomis žvejojantys miškai ir floemai laikomi labiau specializuotais nei neaukštiniai.
Storėjant stiebui, didėja ir kambio sluoksnių perimetras. Tai įmanoma dėl kambinių ląstelių dalijimosi radialine kryptimi.
Tarp kambinių ląstelių yra izoliuotas vidurinis pradinis sluoksnis. Dėl dalijimosi jos ląstelės sudaro darinius vidinėje, o vėliau ir viduje kryptis į išorę. Per tą patį laikotarpį 2-3 ląstelės nusėda į vidų, o tik viena ląstelė nusėda išorine kryptimi. Dėl to susidaro daugiau medienos nei bastos.
Spindulinės ląstelės (inicialai) formuoja šerdies parenchiminius spindulius: pirminius, kurie tęsiasi nuo šerdies iki žievės; o antriniai šerdies spinduliai yra trumpesni, nes atsiranda iš vėliau atsiradusių spindulių ląstelių.
Šerdies sijos gali būti vienos eilės ir kelių eilių; vienalytės (t. y. susidedančios iš tų pačių ląstelių) ir nevienalytės (spindulyje yra skirtingos struktūros ląstelės).
Šerdies spindulių reikšmė : tai maistinių medžiagų (krakmolo, lipidų) saugojimo vieta, kuri atlieka tam tikrą vaidmenį ramybės periodu (pavyzdžiui, žiemą).
Šerdies spinduliai užtikrina radialinį vandens ir mineralinių druskų perkėlimą iš ksilemo išilgai apoplasto į kambį ir antrinį floemą ir priešinga kryptimi išilgai maistinių medžiagų simpplasto.
Ir galiausiai per tarpląstelines erdves vyksta dujų mainai.
Mediena
Medieną formuoja kambis ir ji auga išcentrine kryptimi. Nes mūsų zonoje kambis veikia periodiškai: pradeda funkcionuoti ankstyvą pavasarį ir nustoja veikti vasaros pabaigoje, tada medienoje matomas koncentrinis sluoksniavimasis. Pavasarį, vegetacijos pradžioje, susidaro ankstyva mediena. Jo indai platūs, plonasieniai, galintys pernešti nemažą kiekį vandens. Vanduo reikalingas augimui inicijuoti, ypač norint padidinti naujų ląstelių dydį (pvz., besivystančių lapų ląsteles). Vėlyvoje medienoje yra mažiau laivų. Jie yra siauresni ir storesni. Perėjimas nuo ankstyvos iki vėlyvos tų pačių metų medienos gali būti laipsniškas. Tačiau vienerių metų vėlyvos medienos riba su ankstyvąja mediena kitais metais visada aštrus ir aiškiai matomas skerspjūvyje. Dėl to susidaro metinis augimas (metinis žiedas arba metinis sluoksnis).
Pagal indų išsidėstymą metiniame (metiniame) augime išskiriamos šios medienos rūšys.
Jei indai daugiausia yra ankstyvojoje medienoje, o vėlyvoje medienoje jų nėra arba dedami tik nedideli indai, pvz., ąžuolas, uosis, tai yra žiedo formos mediena.
Jei indai yra tolygiai pasiskirstę per visą metinį augimą, nors vėlyvoje medienoje jų skersmuo mažėja, tai yra išsibarsčiusios kraujagyslinės medienos. Aptinkama berže, kleve, tuopoje.
Taip pat yra pereinamųjų formų tarp šių medienos rūšių.
Kadangi mediena atlieka laidžias, mechanines ir saugojimo funkcijas, ji, be ksilemo elementų, taip pat apima mechaninį audinį sklerenchimos pavidalu ir saugojimo audinį pagrindinės parenchimos pavidalu. Visų anatominių elementų struktūra, jų skaičius ir išsidėstymas suteikia skirtingų augalų medienai specifinių bruožų, būdingų konkrečiai taksonominei grupei.
Pavyzdžiui, augalai, kurie yra labiau pažengę evoliucijos požiūriu, turi specializuotą mechaninį audinį (sklerenchimą). Evoliuciškai primityvūs dviskilčiai (magnolijos) neturi medienos plaušų. Mechaninį vaidmenį šiuo atveju atlieka tracheidai.
Kai kurių augalų medienoje yra pereinamųjų formų nuo tracheidų iki libriforminių (libriforminiai yra specializuoti mechaniniai medienos elementai, susidedantys iš prosenchiminių ląstelių, kurių galai yra nukreipti storais lignifikuotais apvalkalais). Taip pat yra kloisono libriformas. Jo originali verpstės formos ląstelė su storomis vertikaliomis membranomis yra padalinta į atskiras trumpalaikes ląsteles. Tokie histologiniai elementai morfologiškai ir funkciškai artimi sumedėjusiai parenchimai.
Medienos parenchima atlieka saugojimo ir iš dalies laidų vaidmenį. Parenchimos ląstelės yra gyvos. Medienos parenchima dedama į horizontalias ir vertikalias eilutes. Horizontalios eilutės sudaro pagrindinius spindulius. Mes jau apsvarstėme jų savybes. Vertikalios eilės sudaro pačią sumedėjusią parenchimą.
Jei medienos parenchima yra išsklaidyta per visą metinį augimą, tai difuzinė parenchima. Būdinga liepai, ąžuolui, kriaušei.
Jei medienos parenchima yra aplink indus, tai paratracėjinė parenchima. Uosis, klevo mediena.
Ant išorinės ribos yra metinis augimas terminalo parenchima. Pavyzdys: gluosnio, maumedžio, magnolijos mediena.
Jo struktūros ypatumas yra kraujagyslių nebuvimas. Trachėjos elementai yra neperforuoti ir juos vaizduoja tracheidės, atliekančios ir laidų, ir mechaninį vaidmenį.
Spygliuočių tracheidoms būdingos didelės ribojamos poros su toru. Tracheidos yra ilgos su smailiais galais. Ankstyvosios tracheidos yra didelės, plonasienės. Vėliau tracheidės yra radialiai suplotos, storasienės.
Matome, kad gimnasėklių ir dviskilčių mediena turi didelių struktūrinių skirtumų. Todėl jis skirstomas į dvi pagrindines grupes. Gimnosėklių mediena priskiriama minkštai, o dviskilčių – kietajai. Šie terminai „kietas“ ir „minkštas“ nebūtinai apsiriboja tankio ir kietumo laipsniu. Skirtumai, kaip jau minėjome, yra susiję su struktūrinėmis savybėmis.
Pasvirusiuose arba išlenktuose kamienuose atsiranda reaktyvioji mediena. Jo susidarymą lemia šakų ir kamienų polinkis atsispirti apkrovoms, atsirandančioms joms pasvirus, t.y. jo formavimas yra susijęs su šių augalo dalių tiesinimo procesu.
Svarbiausi veiksniai, turintys įtakos reaktyviosios medienos vystymuisi, yra gravitacinės jėgos ir endogeninių augimo stimuliatorių pasiskirstymas.
Reaktyvioji mediena skiriasi nuo įprastos medienos tiek anatomiškai, tiek chemiškai. Reaktyviosios medienos ląstelės labiau lignifikuojamos ir storėja sienelės. Spygliuočiuose jis yra tankesnis ir tamsesnis nei aplinkiniai audiniai. Jo tracheidai yra trumpesni nei įprastos medienos.
Kamieninės medienos radioaktyvumas mažėja nuo išorinių sluoksnių link šerdies. Šiek tiek padidėja medienos užterštumas nuo kamieno pagrindo iki viršaus.
Pakankamai didelis radioaktyviųjų medžiagų kiekis žievėje ir ūgliuose rodo jų išorinį oro užterštumą.
Bark
Šis terminas apima visus audinius, esančius už kambio ribų. Tai jau antrinė žievė, nes. susidaro antrinė meristema – kambis (priešingai nei pirminė, kuri susidaro diferencijuojant pirminės meristemos ląsteles).
Antrinės žievės sudėtis apima antrinę floemą, sklerenchimą (šūnų pluoštus ir akmenines ląsteles) ir pagrindinę parenchimą. Mechaninių elementų rinkinys vadinamas kietas kotas, laidūs elementai ir pagrindinė parenchima - minkštas bastas.
Sekrecinės ląstelės ir dervos kanalai yra antrinėje žievėje. Pagrindinė antrinės žievės funkcija yra plastikinių medžiagų laidumas, taip pat apsaugos funkcija.
Antrinė floema sudaro daug mažesnę kamieno tūrio dalį nei antrinė ksilema. Taip yra dėl to, kad, pirma, kambis sudaro mažiau floemo nei ksilemas, antra, sena, neveikianti floema palaipsniui susmulkinama.
Dėl ašinio organo perimetro padidėjimo parenchiminiai spinduliai plečiasi ir primena trikampį, kurio viršūnė nukreipta į kambį. Tokia parenchima vadinama išsiplėtusia (lot. dilatatio – išsiplėtimas). Toks parenchimos išsidėstymas neleidžia stiebui plyšti, kai stiebas auga.
Spygliuočiuose antrinė floema yra paprastesnės struktūros. Gimnosėklių basoje tolygiai pasiskirsto histologiniai elementai ir, žinoma, yra dervos kanalų.
Visi audiniai, esantys už felogeno ribų (tai reiškia vidinį kamščio kambį), nustoja gauti vandens ir mineralų, nes kamštienos ląstelės tampa suberinizuotos. Visos šios peridermos su žievės liekanomis (negyvomis parenchimo ląstelėmis ir floemu) sudaro išorinę (išorinę) žievę. Gyvoji žievės dalis tarp kambio ir vidinio felogeno sluoksnio vadinama vidine žieve.
Taigi, remdamiesi žiniomis iš temos „Audiniai“, ištyrėme stiebo, kaip ašinio ūglio organo, anatominę struktūrą.
Dabar apsvarstykite šoninio ūglio organo - lapo - struktūrą.
LAPAS
Lapas yra pagrindinis aukštesniųjų augalų fotosintezės organas. Lapo struktūra ir jo funkcijos yra glaudžiai tarpusavyje susijusios.
Iš visos fotosintezės reakcijų lygties:
CO 2 + H 2 O chl-l → hv (CH 2 O) n + O 2
galime daryti išvadą, kad 1) lapams reikalingas CO 2 ir vandens šaltinis; 2) lapai turi būti pritaikyti absorbcijai saulės energija juose turi būti chlorofilo; 3) O 2 išsiskirs kaip vienas iš reakcijos produktų; 4) angliavandeniai turi būti laikomi sandėlyje arba transportuojami į kitas augalo dalis. Sąrašas yra labai specializuota įstaiga, atitinkanti visus šiuos reikalavimus.
Visa lapo, kaip vegetatyvinio organo, raida ėjo prisitaikymo prie geriausio šviesos panaudojimo vystymosi keliu.
Pagal mikrofilinę evoliucijos liniją lapas laikomas archajiškų rinofito tipo augalų telomos atauga ir išlaiko ašinio organo struktūrinę struktūrą.
Pagal makrofilinę evoliucijos liniją, lapas atsirado dėl išlyginimo, telomų išsidėstymo vienoje plokštumoje ir vėlesnio jų susiliejimo į vieną visumą. Tuo pačiu metu buvo prarastas ilgalaikio viršūninio augimo ir šakojimosi gebėjimas. Kokios morfologinės ir anatominės lapo ypatybės, atsiradusios dėl ilgos evoliucijos?
Suaugęs lapas paprastai susideda iš lapo mentės ir lapkočio. Lapkočiai- siaura į stiebą panaši lapo dalis tarp lapo mentės ir ūglio mazgo, kurios pagalba lapas orientuojamas erdvėje ir išdėstomas palankiausiai šviesos atžvilgiu. Per lapkotį jungiasi lapo mentė ir stiebas. Žiedlapyje gerai išsivystę laidūs ir mechaniniai audiniai. Nukritus lapams, lapkotyje susidaro atskiriamasis sluoksnis.
Žemiausia lapo dalis, sujungta su stiebu, vadinama lakštinis pagrindas. Lapo pagrindas gali būti įvairių formų. Dažniausiai tai atrodo kaip nedidelis sustorėjimas ir vadinamas lapų padas. Dažniau lapo apačioje yra skirtingos formos ir porinių šoninių ataugų dydis – stipuliai. Stipulės gali būti lapo formos ir tarnauti kaip papildomas asimiliuojantis paviršius. Stipulės gali būti adatos formos ir veikti kaip augalų apsauga.
Yra pleiskanojančių, membraninių dėmių. Evoliucine prasme stipulių mažėja, o evoliuciškai jaunuose augaluose jie anksti nukrenta arba jų visai nėra.
Kai kurių javų salieruose (skėtiniuose) lapo pagrindas išauga ir suformuoja uždarą arba atvirą vamzdelį – lapo apvalkalą. Lapų apvalkalas prisideda prie ilgalaikio tarpkalnio meristemos išsaugojimo ir yra papildoma ūglio atrama.
Pagrindinė lapo dalis yra lapo ašmenys. Dėl netolygaus viršutinės ir apatinės lapo pusių apšvietimo plokštelė turi dorsoventralinę struktūrą, t.y. lapo viršutinės pusės struktūra, ventralinė, skiriasi nuo apatinės, nugarinės, struktūros. Viršutinė lapo pusė vadinama ventraline, nes. inkstuose ši pusė yra vidinė ir pasukta link ūglio ašies. Apatinė, nugarinė, lapo pusė pumpuruose yra pasukta į išorę nuo augalo. Taigi, dorsoventralinės simetrijos lapas yra dvišalis arba dvipusis arba dvipusis. būdingas dviskilčiams.
Lapas, turintis radialinę simetriją, yra lygiagretus. Tokie sustorėję, cilindro formos lapai randami Crassulaceae. Vienpusiai, arba pavieniai vienpusiai, lapai būdingi javams, svogūnams.
Apsvarstykite anatominė lapų mentės struktūra.
Kaip ir šaknis ir stiebas, lapas susideda iš vientisų, laidžių ir pagrindinių audinių. Nes lapas dažniausiai neturi antrinio augimo (išskyrus nedidelį lapkočių ir stambių gyslų augimą), jis išlaiko epidermį kaip vientisą audinį.
Stomatai dažniausiai randami apatinėje lapo pusėje. Dviskilčių lapų stomatai yra išsibarstę po visą lapo paviršių nematoma tvarka. Vienaskilčiuose ir spygliuočių lapuose jie pasiskirstę eilėmis, lygiagrečiomis išilginei lapo ašiai. Kai kurių javų epidermyje yra motorinių arba motorinių ląstelių. Jie yra didesni nei įprastos epidermio ląstelės. Vakuolė užima beveik visą ląstelę. Praradus turgorą, šios ląstelės susitraukia ir prisideda prie lapo susilankstymo ar susiraityimo.
Didžioji dalis pagrindinio lapo ašmenų audinio yra mezofile. Mezofilas yra diferencijuotas. Palisade mezofilas yra viršutinėje plokštelės pusėje, kempinė - apačioje. Tokios struktūros lapai yra dviveidžiai arba dorsoventraliniai.
Jei palisadinis mezofilas yra abiejose lapo pusėse, lapas yra vienaveidis arba izoliuotas.
Javų mezofile nėra ryškios diferenciacijos į palisadą ir kempinę chlorenchimą. Mezofilo ląstelės išsidėsčiusios radialiai aplink ryšulius.
Laidieji lapų ryšuliai vadinami gyslomis, o išsišakojusi gyslų sistema – vadinama vėdinimas. Kraujagyslinių ryšulių išdėstymo lapų ašmenyse ypatybės lemia skirtingus venų tipus, kurie gali būti dvilypiai, lygiagrečiai, lenkti, delniniai ir plunksniniai.
Priklausomai nuo to, kaip tarpusavyje jungiasi aukštesnių išsišakojusių kategorijų gyslos, išskiriamos atviros (dichotominės) ir uždaros venos. Esant dichotominei venacijai, gyslos šakojasi dichotomiškai, nukrypsta ūmiu kampu ir pasiekia lapo ašmenų kraštą, niekur nesusiliedamos su gretimomis gyslomis. Esant uždarai ventiliacijai, mažos venos dėl anastomozių (susikryžiavimo) sudaro tinklą, prasiskverbiantį per visą plokštelę arba didžiąją jos dalį.
Lapai su delniniu ir plunksniniu gyslumu, dažniausiai dviskilčiai, turi vidutinę, didžiausią gyslą. Veną sudaro pirminis ksilemas ir pirminis floemas, atsirandantis iš prokambiumo ir sujungtų į papildomus ryšulius. Tarp ksilemo ir floemo dažnai susidaro kambis, tačiau jis neveikia. Todėl ksilemas ir floemas yra pirminiai.
Ryšuliai lape sudaro ištisinę sistemą, sujungtą su stiebo laidžiąja sistema. Todėl ksilemas visada yra orientuotas į morfologiškai viršutinę lapo pusę, o floema visada yra orientuota į morfologiškai apatinę lapo pusę.
Vidurinis šonkaulis yra sujungtas su mažesnėmis šoninėmis venomis. Kiekvienas iš jų yra sujungtas su dar mažesniais, o tie, savo ruožtu, skirstomi į dar mažesnius ir pan. Išsišakojimų skaičius dviskilčiuose svyruoja nuo 2 iki 5 ar daugiau. Mažiausios šakos sudaro ląsteles, kuriose yra uždarytos nedideli plotai mezofilas – areola.
Atviroje dichotominėje venų sistemoje nėra uždarų ląstelių.
Vienaląsčių lapų (su lygiagrečia ventiliacija) išilginės venos yra tarpusavyje sujungtos mažesnėmis gyslomis – komisūriniais ryšuliais, kurie išsidėstę paprastų skersinių tiltelių pavidalu. Laidžių audinių skaičius mažose venose palaipsniui mažėja. Ryšulių galuose ksilemo elementai dažnai tęsiasi didesniu atstumu nei floeminiai. Ksilemas venų galuose dažniausiai susideda iš trumpų trachėjos elementų, floema – iš trumpų siaurų sieto elementų ir didelių palydovinių ląstelių.
Maži kraujagysliniai ryšuliai, lokalizuoti mezofile, yra apsupti vieno ar kelių kompaktiškai susiklosčiusių ląstelių sluoksnių, kurie sudaro pluošto apvalkalą. Ryšulio apvalkalai gali būti parenchiminiai arba sklerenchiminiai. Kai kuriose rūšyse stebima ląstelių sienelių suberinizacija fascikulų apvalkaluose, o tai rodo, kad apvalkalo ląstelės gali veikti kaip endoderma.
Kolenchima ir sklerenchima yra didelėse venose vienoje arba abiejose pusėse. Su didelėmis venomis susijęs audinys pakyla virš lapo paviršiaus ir suformuoja iškilimus. Audinio plotas, uždarytas tarp venų išsikišimų, vadinamas „tarpšonkauline zona“ (lot. Iškyšos pavadinimas „costa“ – šonkaulis).
Rūšių su storais lapais mezofile randama įvairių sklereidų, kurie veikia kaip tarpikliai.
Javų lapams būdingas stiprus sklerenchimos išsivystymas.
Gimnosėklių lapai yra ne tokios įvairios struktūros nei gaubtasėklių lapai. Pagal pušies spyglių pavyzdį apsvarstykite, kaip prisitaikymas toleruoti žemą temperatūrą žiemą ir galimybė gyventi kelerius metus atsispindi anatominėje struktūroje. Adata turi storasienį epidermį su galinga odele ir giliai panirusiomis stomatomis. Kai kurių spygliuočių (Cupressaceae) šoninės ląstelės turi kutikulinį gūbrį, kabantį virš stomatos apsauginių ląstelių. Stomatai yra išdėstyti vertikaliomis eilėmis iš visų spyglių pusių. Po epidermiu yra storasienės, iš dalies sudegintos hipodermio ląstelių membranos. Mezofilas sulankstytas. Jame yra dervos kanalų. Laidūs ryšuliai yra adatos centre. Dažniausiai du, retai vienas. Šie ryšuliai yra sujungti sklerenchiminiu lygintuvu. Ryšulius supa transfuzinis audinys, susidedantis iš tracheidų ir parenchiminių ląstelių. Transfuzinis audinys (lot. transfusio – transfuzija) yra susijęs su vandens ir maisto medžiagų pernešimu tarp laidžių pluoštų ir mezofilo.
Transfuzinį audinį supa storasienė endoderma. Ankstyvosiose vystymosi stadijose endodermoje dažnai yra kasparinės juostos, o vėlesnėse stadijose – suberinė plokštelė. Subrendusios endodermos ląstelės turi antrines lignified membranas.
Anatominė lapų mentės struktūra atskleidžia didelį plastiškumą. Struktūra kinta priklausomai nuo augalo egzistavimo sąlygų ir lapų išsidėstymo ant augalo.
Jau pažymėjome vieną iš šiuolaikinės botanikos raidos krypčių – ekologinę augalų anatomiją – ir teigėme, kad būtent gyvasis ląstelės turinys – protoplastas – pirmiausia reaguoja į aplinkos sąlygų pokyčius. Taigi šiaurinių augalų ląstelės yra mažiau vakuoliuotos ir turi didelius chloroplastus. Stromoje yra daug krakmolo grūdelių. Chloroplastų tilakoidų sistema yra mažiau išsivysčiusi nei vidutinių platumų augaluose. Šiauriniuose augaluose mitochondrijų skaičius yra 2-2,5 karto didesnis. EPR ir Golgi aparatai yra labiau išvystyti. Tokios šiaurinių augalų mezofilo ląstelių submikroskopinės struktūros ypatybės apibūdinamos kaip svarbūs prisitaikymo bruožai, prisidedantys prie augalų išlikimo atšiauriomis klimato sąlygomis.
Augaluose, prisitaikiusiuose prie drėgmės trūkumo, pastebimos šaltos, storos ląstelių membranos, ypač epidermyje. Stipriai sustorėjusios stambiųjų ląstelių epidermio išorinės membranos užima pusę ląstelės ertmės. Virš epidermio yra storas odelių sluoksnis. Iš paviršiaus odelė pasidengia storomis vaško apnašomis, kurios lemia melsvą lapų atspalvį.
Stomatos yra panardintos į įdubas, kurias išstumia epidermio plaukeliai.
Sultingi augalai turi vandens kaupimo audinį.
Įdomūs pritaikymai drėgmei sulaikyti minkštųjų quinoa lapuose (Afrika). Lapas iš viršutinės ir apatinės pusės yra padengtas ištisiniu daugiaeilių pūslelių sluoksniu. Šio sluoksnio storis gerokai viršija mezofilo storį ir apsaugo gyvus audinius nuo išsausėjimo. Plaukai gali sulaikyti oro drėgmę naktį ir pagerinti šiurkštumą vandens režimas augalai.
Mūsų besidriekiančios miško zonos kvinojos lapai ir stiebas yra padengti daugybe burbuliukų pavidalo plaukelių, sukuriančių miltelinės dangos įspūdį, tačiau jie nesudaro tokio tankaus kelių eilių sluoksnio, kaip minkštojoje kinojoje.
Dviejų rūšių kvinojų lapų palyginimo pavyzdyje matyti, kad panašių taksonominių vienetų sisteminiai bruožai, kai jie apsigyvena skirtingomis ekologinėmis sąlygomis, įgyja nevienodą vystymąsi ir yra prisitaikančio pobūdžio.
Atogrąžų miškų augalų (pavyzdžiui, monsterų) lapai turi skylėtas lapų ašmenis ir ilgus lapkočius. Vandens perteklius visada lengvai nuteka. Stomos yra atviros. Palisade audinys su didelėmis oro ertmėmis.
Be temperatūros ir drėgmės, dar vienas svarbus abiotinis veiksnys – šviesa, į kurią augalas reaguoja su struktūriniais pokyčiais.
Lapai, besivystantys tiesioginiuose saulės spinduliuose, yra mažesni už šešėlinius lapus, bet storesni nei šešėliai dėl stipraus palisadinio mezofilo išsivystymo.
Taigi prisitaikymai prie aplinkos sąlygų, fiksuotų evoliucijos eigoje, yra susiję su kiekybiniais skirtumais, bet nekeičia organizavimo principų.
Lapų ontogeniškumas
Lapai yra išdėstyti mažų gumbų pavidalu, kaip šoninės viršūninės meristemos ataugos. Formuojant lapų gumbą dalyvauja ne tik tunika, bet ir kūno ląstelės. Lapų primordijos auga nuo pradinio išsikišimo į viršų kūgiškos arba adatos formos ataugos pavidalu. Netrukus lapų užuomazgos apgaubia viršūninę meristemą, apsaugodamos ją tiek mechaniškai, tiek nuo šilumos, kurią išskiria kvėpuojant.
Ankstyvas lapų augimas paprastai skirstomas į viršūninį ir kraštinį (ribinį). Pirmasis yra susijęs su primordijų pailgėjimu, antrasis su šoniniu augimu, dėl kurio susidaro dvi plokštelės dalys. Atitinkamai, augančios primordijos turi viršūninę meristemą viršūnėje ir dvi kraštines meristemas ašies šonuose. Primordijų viršūninis augimas nėra ilgas. Plokštelės viršūnės ir kraštų augimą lydi tarpkalnis.
Sudėtiniame lape kraštinės meristemos aktyvumas yra lokalizuotas atskiruose centruose, kurių kiekvienas sudaro atskirą lapelį.
Lapašnio vystymasis yra greitesnis nei lapkočio augimas, kuris vėliau susidaro tarpkalniniu augimu.
Jei tarp tarpkalnio ir kraštinių zonų pagrindo ribos susidaro susiaurėjimas, susidaro lapkočio lapas. Didžiąją lapkočio dalį sudaro tarpkalinė zona. Jei toks susiaurėjimas (ant ribos tarp tarpkalnio ir kraštinių zonų pagrindo) nesusidaro, išsivysto bekojis lapas.
Vienakilčių augalų lapų rudimente beveik nesiformuoja kraštinės meristemos, ilgą laiką būdingas tarpkalinis augimas. Taigi pagrindinė lapo mentės forma yra linijinė.
Skirtinguose plokštelės sluoksniuose esančių ląstelių dalijimosi ir pailgėjimo greičio skirtumai lemia daugybės tarpląstelinių erdvių susidarymą ir tipišką mezofilo struktūrą.
Kitų audinių vystymasis koreliuoja su laidžių audinių vystymusi. Prokambiumas tuo pačiu metu atsiskiria lape ir gretimame augimo kūgio plote, sudarydamas ištisinę sruogą. Jis sudaro pirminį kraujagyslių pluoštą, kuris yra ištisinis ir bendras lapui ir stiebui.
Lapo ontogenezė liudija apie bendrą lapo ir stiebo, kaip vientisos visumos – ūglio – dalių prigimtį.
Nuo pumpuro išsivystymo momento prasideda lapų vystymosi fazė be pumpurų. Tuo pačiu metu lapų paviršius padidėja kelis šimtus, net tūkstančius kartų. Paviršiaus augimas pasiekiamas dalijant daugumą lapų ląstelių ir ištempiant jas į ilgį ir plotį.
Lapų gyvenimo trukmė priklauso nuo genetinių ir klimato veiksnių. Vidutinio klimato lapuočių medžių ir krūmų lapų auginimo laikotarpis yra 4-5 mėnesiai. Spygliuočių augalų lapai gyvena nuo dvejų iki šešerių ar daugiau metų. Tačiau bet kuriuo atveju lapų gyvenimo trukmė yra daug mažesnė nei ašinių augalo organų gyvenimo trukmė.
Welwitschia mirabilis yra kilęs iš pietvakarių Afrikos uolėtų dykumų. Šis žemaūgis medis turi ilgą šaknį, storą ir trumpą kamieną (iki 50 cm aukščio ir iki 1 m storio) ir du didelius lapus, kurie išlieka visą gyvenimą (iki 2000 metų). Lapai siekia 2–3 metrus ir nuolat auga prie pagrindo, miršta viršuje. Beveik vienintelis Velvichia drėgmės šaltinis yra tankus rūkas, kurio drėgmę šis augalas sugeria per daugybę stomatelių abiejose lapo pusėse (22 200 stomatelių 1 cm 2).
Aktyvus lapų atskyrimas nuo šakų, nepažeidžiant jo gyvų audinių, vadinamas abscisija. Sezoninis medžių lapų kritimas yra reakcijos į dienos trukmės pasikeitimą rezultatas. Rudenį krintantys lapai ženkliai sumažina augalo garavimo paviršių, o tai būtina esant fiziologiniam drėgmės trūkumui rudenį ir žiemą. Dėl lapų kritimo sumažėja šakų lūžimo rizika nuo sniego svorio.
Daugumoje lapų atskiriamasis sluoksnis susidaro ontogenezės metu. Šioje zonoje sumažėja mechaninio audinio kiekis. Prieš išsiliejimą trachėjos elementuose susidaro tilai, o sieto elementuose nusėda kaliozė. Abscisijos metu vyksta fermentinis ląstelių membranų sunaikinimas, dėl kurio ląstelės atsiskiria viena nuo kitos. Membranų pakitimai apima vidurinės sluoksnio cementavimo gebėjimo praradimą (iš dalies dėl kalcio pašalinimo iš jos), pačių celiuliozės membranų hidrolizę ir sklerifikuotų trachėjos elementų plyšimą.
Apsauginis sluoksnis arba randas susidaro membranose ir tarpląstelinėse erdvėse nusėdus apsauginėms medžiagoms, tokioms kaip suberinas ar žaizdos guma. At sumedėję augalai apsauginį sluoksnį pakeičia periderma, kuri guli po apsauginiu sluoksniu ir tampa ištisine su likusio stiebo periderma.
Lapų kritimas nebūtinai yra susijęs su ląstelių membranų tirpimo procesu. Metant spyglius nuo eglės, galima pastebėti mechaninį atsiskyrimą be išankstinių cheminių pakitimų.
Nukritus lapams, lieka lapų randai su jų krūvomis. Sijos randai- tai nulūžę kraujagyslių ryšulių galai, kurie nuo lapų pėdsakų nuėjo iki lapo lapkočio prieš jam nukritus.
lapų pėdsakai- tai šakos nuo stiebo laidžiosios sistemos iki lapų. Lapo pėdsakas driekiasi nuo jo susiliejimo su stiebo ryšuliu vietos iki išėjimo į lapą. Viename lape gali būti vienas ar daugiau lapų pėdsakų.
Nes stiebas ir lapas turi filogenetiškai bendrą kilmę, esminio skirtumo tarp lapų pėdsakų ir stiebo ryšulių nėra. Atitinkami terminai turi aprašomąją, topografinę reikšmę.
Mazgo srityje, kur lapo pėdsakas nukrypsta nuo centrinio stiebo laidžiojo cilindro link lapo pagrindo, cilindre susidaro parenchiminė zona - lapo tarpas arba lapo tarpelis. Lapų pėdsakų ir lapų spragų skaičius skiriasi nuo augalo iki augalo.
Pirminis gaubtasėklių mazgų tipas yra toks, kuriame yra trys ar daugiau spragų. Pagrindinė gaubtasėklių evoliucijos kryptis pasižymi spragų skaičiaus mažėjimu.
Spragos, šerdis, tarpfaskulinės zonos, laidžioji sistema ir periciklas, esantys laidžiosios sistemos periferijoje, sudaro vidinę ašinės augalo dalies (stiebo ir šaknies) šerdį arba stela.
Stelų klasifikacija grindžiama laidžių ir nelaidžių audinių tarpusavio išsidėstymu ašiniuose organuose pirminiame vystymosi etape. Paprasčiausio tipo steloje, kuri laikoma filogenetiškai primityviausia, laidus audinys sudaro ištisinę koloną, o ksilemą supa floema. Tai protostele. Tokia stiebo organizacija yra biologiškai netobula, nes kontaktinis paviršius tarp laidžių ir apatinių audinių yra mažas.
Tolesnė stelos evoliucija ėjo tuo keliu, kaip didėjo laidžių audinių kontaktinis paviršius su pagrindiniais. Tai buvo pasiekta dviem būdais. Vienu atveju stela formuoja gilias ataugas link žievės ir yra žvaigždės formos skerspjūvyje. Tai aktinostele.
Antrasis, biologiškai perspektyvesnis vystymosi būdas yra parenchiminės šerdies atsiradimas stelos centre. Tai yra sifonas.
Glaudesnis ryšys tarp laidžių ir pagrindinių audinių atsiranda vystantis lapams. Kuo daugiau lapų, tuo labiau stela yra prisotinta pagrindine parenchima. Stela įgauna tinklo formą (graikiškai tinklas – dictuon). Tai yra diktotele.
Šiuolaikinių žydinčių augalų stela dar labiau parenchimatizuota. Stela, kuri yra laidžių sruogų ir tarpfaskulinių zonų sistema, yra eustela (gr. eu – geras).
Vienakilčių lapų pėdsakai pasiekia stiebo centrą, tada nukrypsta į pakraščius. Todėl ant stiebo skerspjūvio ryšuliai atrodo išsibarstę be jokios tvarkos. Ataktos iš graikų k. netvarkingas. Iš čia ir kilo vienaskilčių stelos pavadinimas – ataktostele. Jai būdinga aukštas laipsnis parenchimatizacija, kai kiekvienas lapo pėdsakas "
Šaknis yra ašinis organas, turintis radialinę simetriją ir augantis tol, kol išsaugoma viršūninė meristema. Šaknis morfologiškai skiriasi nuo stiebo tuo, kad ant jo niekada neatsiranda lapai, o viršūninę meristemą dengia šaknies kepurėlė. Šaknų palikuonių augaluose išsišakojimas ir atsitiktinių pumpurų atsiradimas vyksta endogeniškai (viduje) dėl periciklo (pirminės šoninės meristemos) veiklos.
Šaknies funkcijos:
1) Šaknis sugeria vandenį iš dirvos su joje ištirpusiomis mineralinėmis medžiagomis;
2) atlieka inkaro vaidmenį, fiksuodamas augalą dirvoje;
3) tarnauja kaip maistinių medžiagų talpykla;
4) dalyvauja pirminėje tam tikrų organinių medžiagų sintezėje;
5) šaknų palikuonių augaluose atlieka vegetatyvinio dauginimosi funkciją.
Šaknų klasifikacija. Pagal kilmę šaknys skirstomos į pagrindines, atsitiktines ir šonines. Šaknis, kuris išsivysto iš sėklos gemalinės šaknies, vadinamas pagrindinis; vadinamos šaknys, atsirandančios ant kitų augalų organų (stiebo, lapo, žiedo). priedinis. Atsitiktinių šaknų vaidmuo žolinių gaubtasėklių gyvenime yra didžiulis, nes suaugusių augalų (tiek vienaskilčių, tiek daugelio dviskilčių) šaknų sistemą daugiausia (arba tik) sudaro atsitiktinės šaknys. Papildomų šaknų buvimas bazinėje ūglių dalyje leidžia lengvai padauginti augalus dirbtiniu būdu, dalijant juos į atskirus ūglius arba ūglių grupes su papildomomis šaknimis.
Jaunuose stiebuose atsitiktinių šaknų užuomazgos susidaro endogeniškai - iš tarpfaskulinės parenchimos ląstelių, senuose - iš parenchimos, esančios šalia šerdies spindulių kambio, tai yra, parenchiminės kilmės kambio. Atsitiktinės vienakilčių stiebų šaknys atsiranda ryšulius supančioje parenchimoje.
Šoninėšaknys susidaro ant pagrindinių ir atsitiktinių šaknų. Dėl tolesnio jų šakojimosi atsiranda aukštesnių kategorijų šoninės šaknys. Dažniausiai šakojasi iki ketvirtos ar penktos eilės.
Vadinama visų vieno augalo šaknų visuma šaknų sistema.
pagal kilmę:
pagrindinė šaknų sistema vystosi iš gemalinės šaknies ir yra atstovaujama pagrindinės šaknies (pirmos eilės) su antrosios ir vėlesnių eilės šoninėmis šaknimis. Daugelyje medžių ir krūmų bei vienmečių ir kai kurių daugiamečių žolinių dviskilčių vystosi tik pagrindinė šaknų sistema;
atsitiktinė šaknų sistema vystosi ant stiebų, lapų, kartais ant žiedų. Ši šaknų kilmė vertinama kaip primityvesnė, nes būdinga aukštesnėms sporoms, kurios turi tik atsitiktinių šaknų sistemą. Papildomų šaknų sistema gaubtasėkliuose susidaro iš sėklos, kurioje išsivysto protokormas (embrioninis gumbas), o vėliau ant jo - atsitiktinės šaknys;
mišri šaknų sistema plačiai paplitę tarp dviskilčių ir vienaskilčių. Iš sėklos išaugintame augale pirmiausia išsivysto pagrindinės šaknies sistema, tačiau jos augimas netrunka ilgai – dažnai sustoja iki pirmojo vegetacijos sezono rudens. Iki to laiko atsitiktinių šaknų sistema nuosekliai išsivysto ant pagrindinio ūglio hipokotilo, epikotilo ir vėlesnių metamerų, o vėliau ir šoninių ūglių bazinėje dalyje. Priklausomai nuo augalų rūšies, jie inicijuojami ir vystomi tam tikrose metamerų dalyse (mazguose, po ir virš mazgų, tarpmazguose) arba per visą ilgį.
Šaknų sistemų klasifikacija informuoti.
Pagrindinė šaknų sistema vadinama kertinis jei pagrindinė šaknis ilgiu ir storiu pastebimai viršija šonines;
Panašaus dydžio pagrindinės ir šoninės šaknys, šaknų sistema pluoštinis. Taip pat gali būti mišri šaknų sistema kertinis, jei pagrindinė šaknis yra daug didesnė už kitas, pluoštinis jeigu visos šaknys santykinai vienodo dydžio. Tie patys terminai galioja ir atsitiktinių šaknų sistemai.
Toje pačioje šaknų sistemoje šaknys dažnai atlieka skirtingas funkcijas. Yra kaulinės šaknys (atraminės, tvirtos, su išsivysčiusiais mechaniniais audiniais), augančios šaknys (greitai augančios, bet mažai šakojasi), čiulptančios (plonos, trumpaamžės, intensyviai šakojasi).



































- Dydis: 2 MB
- Skaidrių skaičius: 36
Prezentacijos aprašymas ROOT AND ROOT SYSTEMS 1. Funkcijos ir evoliucinės skaidrės
ŠAKNŲ IR ŠAKNŲ SISTEMOS 1. Šaknies funkcijos ir evoliucinė kilmė. 2. Pirminė šaknies struktūra. 3. Antriniai šaknų pokyčiai. 4. Šoninių ir atsitiktinių šaknų formavimasis. šaknų sistemos. 5. Šaknų specializacija ir modifikavimas.
Šaknis yra ašinis organas, turintis radialinę simetriją ir neribotą ilgį augantis dėl viršūninės meristemos veiklos. Lapai niekada neatsiranda ant šaknies, o viršūninė meristema visada yra padengta kepure. Pagrindinė šaknies funkcija yra vandens ir mineralų įsisavinimas, tai yra, augalo aprūpinimas dirvožemiu. Be minėtos pagrindinės funkcijos, šaknys atlieka ir kitas funkcijas: stiprina augalą dirvožemyje, leidžia augti vertikaliai ir šaudyti; šaknyse vyksta antrinė įvairių medžiagų (amino rūgščių, alkaloidų, fitohormonų ir kt.) sintezė; atsarginės medžiagos gali nusėsti į šaknis; šaknys sąveikauja su kitų augalų šaknimis, dirvožemio mikroorganizmais ir grybais.
Šaknys atsirado iš rinofitų telomų, išplitusių dirvos paviršiuje. Evoliucijos eigoje kai kurios šių telomų šakos pradėjo gilėtis į dirvą ir davė šaknis.
Šaknys prisitaiko prie tobulesnės dirvos mitybos. Šaknų atsiradimą lydėjo gilus visos jų struktūros pertvarkymas. Jie turi specializuotus audinius. Medžiagų sugėrimo iš dirvožemio funkciją pradėjo atlikti jaunos šaknų galūnės. Jie išlaiko gyvas ląsteles paviršiuje. Šios ląstelės sudarė funkciškai svarbiausią šaknies audinį – rizodermą.
Medžiagų sugėrimo iš dirvožemio funkciją pradėjo atlikti jaunos šaknų galūnės. Jie išlaiko gyvas ląsteles paviršiuje. Šios ląstelės sudarė funkciškai svarbiausią šaknies audinį – rizodermą. Toliau evoliucijos procese absorbcinis šaknies paviršius didėjo dėl trijų veiksnių: 1) gausaus šakojimosi ir daugybės sugeriamųjų galūnių susidarymo; 2) nuolatinis šaknų augimas ir siurbimo galūnių judėjimas į naujas dirvožemio vietas; 3) šaknų plaukelių susidarymas.
Kadangi šaknis auga tankiame dirvožemyje, jos viršūninė meristema turi būti apsaugota. Viršūninę meristemą nuo pažeidimų apsaugojo atsiradęs šaknies dangtelis. Šaknų atsiradimą lėmė padidėjęs klimato sausumas. Dėl sausesnio klimato sausumos augalai prisitvirtino prie substrato ir iš jo pasisavina vandenį bei maistines medžiagas. Tačiau evoliucijos eigoje skirtingų augalų rūšių šaknų struktūra kito mažiau nei stiebo. Taip yra dėl to, kad sąlygos dirvožemio aplinkoje yra stabilesnės nei ore. Todėl šaknis laikomas „konservatyvesniu“ organu, nors atsirado daug vėliau nei pabėgimas. Šaknų formavimasis yra svarbi augalų aromorfozė. Jo dėka augalai sugebėjo įvaldyti sausesnes dirvas ir suformuoti didelius aukštyn kylančius ūglius.
Šaknies dangtelio amiloplastų reakcija į gravitaciją. Judantys statolitai vaidina svarbus vaidmuo kuriant fitohormonų gradientus, kurie užtikrina vertikalų šaknų augimą.
Kviečių (Triticum aestivum) sodinuko šaknies struktūra: A - šaknies struktūros diagrama; B – rizoderminių ir egzoderminių ląstelių diferenciacija. 1 - laidumo zona, 2 - siurbimo zona, 3 - tempimo zona, 4 - dalijimosi zona, 5 - šaknų plaukai, 6 - šaknų dangtelis.
Šaknies skerspjūvis (a - vienaskiltis, b - dviskiltis augalas)
Pirminė žievė kyla iš periblemos. Jo pagrindinę masę sudaro gyvos parenchiminės ląstelės su plonomis membranomis. Tarp jų susidaro tarpląstelinių erdvių sistema, pailgėjusi palei šaknies ašį. Dujos (CO 2) cirkuliuoja per tarpląstelines erdves. Dujos būtinos norint palaikyti intensyvų medžiagų apykaitą žievės ir rizodermos ląstelėse. Energetinė medžiagų apykaita žievės ląstelėse būtina, kad būtų galima atlikti daugybę svarbių funkcijų: 1) žievės ląstelės aprūpina šakniastiebį plastikinėmis medžiagomis ir pačios dalyvauja medžiagų įsisavinime ir laidumoje; 2) žievėje sintetinamos įvairios medžiagos, kurios vėliau pernešamos į kitus audinius; 3) žievės ląstelėse kaupiasi atsarginės medžiagos; 4) žievėje dažnai yra grybų hifai, kurie formuoja mikorizę.
Endodermos ląstelės pereina tris vystymosi etapus. Absorbcijos zonoje endoderma yra pirmoje stadijoje. Jo ląstelių radialinių sienelių viduryje susidaro kasparinės juostos. Kasparinės juostos blokuoja medžiagų judėjimą per ląstelių membranas, ty išilgai apoplasto. Antrasis etapas gali būti stebimas šoninių šaknų zonoje. Tuo pačiu metu vidinėje ląstelės membranos pusėje atsiranda plona suberino plokštelė. Tačiau endoderma vis dar gali laisvai praeiti tirpalus, nes joje yra atskirų ląstelių su plonomis sienelėmis. Trečiasis endodermos vystymosi etapas gali būti stebimas vienaskilčių šaknų srityje. Jo ląstelių vidinės ir radialinės sienelės yra labai sustorėjusios. Skersinėse pjūviuose šios ląstelės turi pasagos formą. Patikrinimo punktų nėra. Storasienė endoderma apsaugo laidžius audinius ir padidina šaknų stiprumą.
Kasparijos diržas yra vandeniui atsparus barjeras, kuris verčia vandenį palikti apoplastą ir per endodermos ląstelių membranas veržtis į simpplastą.
Vandens pritekėjimas iš dirvožemio į šaknį: vanduo gali judėti apoplastu ir simpplastu, kol pasiekia endodermą. Tolesnis judėjimas išilgai apoplasto neįmanomas.
Skirtingi šaknies centrinio cilindro struktūros tipai (pirminė struktūra): A-diarchija, B-triarchija, C-tetraarchija, G-poliarchija. A-B tipai būdinga dviskilčiams, G – daugeliui vienakilčių. 1 - pirminės žievės sritis, 2 - pirminė floema, 3 - pirminė ksilema.
Galima išskirti 4 šaknies perėjimo iš pirminės į antrinę struktūrą stadijas: 1) kambio atsiradimas tarp pirminio floemo ir ksilemo sričių; 2) Felogeno susidarymas periciklu; 3) pirminės plutos išsiliejimas; 4) laidių audinių radialinio išsidėstymo pakeitimas kolateraliniais.
Perėjimas iš pirminės į antrinę šaknų struktūrą , 11 - antrinė žievė, 12 - felogenas, 13 - flemas.
Žirnio šaknies laidžių audinių pirminės diferenciacijos schema 1 - ksilemo įcentrinė diferenciacija, 2 - epidermis, 3 - pirminė žievė, 4 - endodermis, 5 - pirmieji diferencijuoti ksilemo elementai, 6 - nediferencijuoti ksilemo elementai, 7 - pirmieji diferencijuoti floemo elementai , 9 - viršūninė meristema, 10 šaknų kepurė.
Antriniai vienakilčių šaknų pokyčiai. Didžioji dauguma vienaskilčių augalų pirminę šaknų struktūrą išlaiko iki savo gyvenimo pabaigos. Tačiau daugelis šaknies elementų lignifikuojami. Sumedėjusių vienakilčių (palmių, dracenų, jukų) šaknies žievėje iš parenchimos ląstelių arba iš periciklo susidaro meristeminis sluoksnis. Iš jo susidaro uždarų laidžių ryšulių eilės. Po šios laidžių ryšulių serijos pirminės žievės parenchimo periferinėje dalyje atsiranda naujas edukacinio audinio sluoksnis. Šis meristemų sluoksnis sukuria naują kraujagyslių pluoštų seriją. Taigi šaknis sustorėja.
Atsitiktinės šaknys atsiranda ant įvairių augalų organų – ant stiebų, lapų ir šaknų. Atsitiktinės šaknys, atsiradusios ant stiebo, vadinamos stiebo šaknimis, o atsiradusios ant šaknies – šaknies šaknimis. Šoninės ir atsitiktinės šaknys yra endogeninės kilmės, tai yra, jos yra išdėstytos vidiniuose audiniuose.
Šoninės šaknies iniciacija prasideda nuo periciklo ląstelių dalijimosi. Šiuo atveju stelos paviršiuje susidaro meristeminis gumbas. Po meristeminio tuberkulio ląstelių dalijimosi serijos atsiranda šoninė šaknis. Jis turi savo viršūninę meristemą ir apvalkalą. Šoninės šaknies pumpuras išauga, prasiskverbia pro pirminę motininės šaknies žievę ir juda į išorę. Paprastai šoninės šaknys kyla prieš ksilemo elementus. Todėl jie yra išdėstyti taisyklingomis išilginėmis eilėmis išilgai šaknies. Jie atsiranda absorbcijos zonoje arba šiek tiek aukščiau. Šoninės šaknys klojasi akropetiškai, t.y. nuo šaknies pagrindo iki jos viršūnės.
Atsitiktinės šaknys dažniausiai dedamos į meristeminę veiklą galinčius audinius: periciklą, kambį, felogeną. Endogeninis šoninių (ir atsitiktinių) šaknų formavimasis turi adaptacinę reikšmę. Jei šakojimasis vyktų viršūnėje, tada šaknies įsigalėjimas dirvoje būtų sunkus.
Dichotominis šakų išsišakojimas skroblinių samanų (Lycopodium clavatum) 1 šaknų sistemoje – ploniausių šaknų izotominis dichotominis išsišakojimas
Mikorizė: A – ektotrofinė ąžuolinė mikorizė, B, C – endotrofinė orchidėjų mikorizė. Mazgeliai ant lubinų šaknų