Хажуугийн үндэс үүсэх. Үндэс, түүний үүрэг. Үндэс ба үндэс системийн төрлүүд
Үндэс нь ургамлын газар доорх тэнхлэгийн элемент бөгөөд тэдгээрийн хамгийн чухал хэсэг, ургамлын гол эрхтэн юм. Үндэсний ачаар ургамал нь хөрсөнд тогтож, амьдралынхаа туршид тэнд хадгалагдаж, түүнд агуулагдах ус, эрдэс бодис, шим тэжээлээр хангадаг. Төрөл бүрийн үндэс, төрөл зүйл байдаг. Тэд тус бүр өөрийн гэсэн онцлог шинж чанартай байдаг. Энэ нийтлэлд бид одоо байгаа үндэс, үндэс системийн төрлүүдийг авч үзэх болно. Мөн бид тэдний онцлог шинж чанаруудтай танилцах болно.
Ямар төрлийн үндэс байдаг вэ?
Стандарт үндэс нь судалтай эсвэл нарийн цилиндр хэлбэртэй байдаг. Олон ургамалд үндсэн (үндсэн) үндэсээс гадна бусад төрлийн үндэс үүсдэг - хажуугийн болон нэмэлт. Тэд юу болохыг илүү нарийвчлан авч үзье.
үндсэн үндэс
Энэ ургамлын эрхтэн нь үрийн үр хөврөлийн үндэсээс үүсдэг. Үргэлж нэг үндсэн үндэс байдаг (бусад төрлийн ургамлын үндэс нь ихэвчлэн олон тооны байдаг). Энэ нь амьдралынхаа туршид ургамалд үлддэг.
Үндэс нь эерэг геотропизмаар тодорхойлогддог, өөрөөр хэлбэл таталцлын нөлөөгөөр субстрат руу босоо чиглэлд гүнзгийрдэг.
адал явдалт үндэс
Адал явдал гэж нэрлэдэг ургамлын үндэс нь тэдний бусад эрхтэн дээр үүсдэг. Эдгээр эрхтнүүд нь иш, навч, найлзуурууд гэх мэт байж болно. Жишээ нь, үр тариа нь үрийн үр хөврөлийн ишэнд байрладаг анхдагч adventitious үндэс гэж нэрлэгддэг. Тэд үрийн соёололтын явцад үндсэн үндэстэй бараг нэгэн зэрэг хөгждөг.
Мөн навчны шинэ төрлийн үндэс (навчны үндэс ургасны үр дүнд үүссэн), иш эсвэл зангилаа (үндэслэг иш, газар дээрх эсвэл газар доорх ишний зангилаанаас үүссэн) гэх мэт байдаг. Хүчтэй үндэс нь доод зангилаанд үүсдэг. агаарын (эсвэл дэмжих).
Нэмэлт үндэс гарч ирэх нь ургамлын ургамлын үржих чадварыг тодорхойлдог.
Хажуугийн үндэс
Хажуу талыг хажуугийн мөчир хэлбэрээр үүссэн үндэс гэж нэрлэдэг. Тэд үндсэн болон нэмэлт үндэс дээр хоёуланд нь үүсч болно. Нэмж дурдахад тэдгээр нь хажуугаас салаалж болох бөгөөд үүний үр дүнд дээд зэрэглэлийн хажуугийн үндэс (эхний, хоёр, гуравдугаар) үүсдэг.
Хажуугийн том эрхтнүүд нь хөндлөн геотропизмаар тодорхойлогддог, өөрөөр хэлбэл тэдгээрийн өсөлт нь бараг хэвтээ байрлалд эсвэл хөрсний гадаргуутай өнцөгт үүсдэг.
Үндэс систем гэж юу вэ?
Үндэс системийг нэг ургамалд байдаг бүх төрлийн үндэс (өөрөөр хэлбэл тэдгээрийн нийт) гэж нэрлэдэг. Үндсэн, хажуугийн болон нэмэлт үндэсийн өсөлтийн харьцаанаас хамааран түүний төрөл, шинж чанарыг тодорхойлно.
Үндэс системийн төрлүүд
Хэрэв үндсэн үндэс нь маш сайн хөгжсөн бөгөөд өөр зүйлийн үндэс дунд мэдэгдэхүйц байвал энэ нь ургамал саваа системтэй гэсэн үг юм. Энэ нь ихэвчлэн хоёр талт ургамлаас олддог.
Энэ төрлийн үндэс систем нь хөрсөнд гүн соёололтоор тодорхойлогддог. Жишээлбэл, зарим өвсний үндэс нь 10-12 метрийн гүнд (аргас, царгас) нэвтэрч болно. Модны үндэс нэвтрэх гүн нь зарим тохиолдолд 20 м хүрч болно.
Хэрэв гэнэтийн үндэс нь илүү тод, олон тоогоор хөгжиж, гол нь удаан өсөлтөөр тодорхойлогддог бол үндэс систем үүсдэг бөгөөд үүнийг фиброз гэж нэрлэдэг.

Дүрмээр бол зарим өвслөг ургамал нь ийм системээр тодорхойлогддог. Шилэн системийн үндэс нь саваа системийн үндэс шиг гүн нэвтэрдэггүй ч зэргэлдээх хөрсний хэсгүүдийг илүү сайн сүлждэг. Элбэг ширхэгтэй нимгэн үндэс үүсгэдэг олон сул бутлаг, үндэслэг ишт өвсийг жалга, энгэрийн хөрс гэх мэтийг засахад өргөн ашигладаг. Шилдэг ширэгт өвсүүдэд саравчгүй гал, шаргал болон бусад орно.

өөрчлөгдсөн үндэс
Дээр дурдсан ердийн зүйлээс гадна бусад төрлийн үндэс, үндэс системүүд байдаг. Тэднийг өөрчлөгдсөн гэж нэрлэдэг.
хадгалах үндэс
Нөөцөд үндэс үр тариа, үндэс булцуу орно.
Үндэс үр тариа нь шим тэжээлийн бодис хуримтлагдсанаас үндсэн үндэс нь өтгөрдөг. Мөн ишний доод хэсэг нь эх үр тариа үүсгэхэд оролцдог. Ихэнхдээ хадгалалтын суурь эдээс бүрддэг. Үндэс үр тарианы жишээ бол яншуй, улаан лууван, лууван, манжин гэх мэт.

Хэрэв өтгөрүүлсэн хадгалалтын үндэс нь хажуугийн ба нэмэлт үндэс байвал тэдгээрийг үндэс булцуу (боргоцой) гэж нэрлэдэг. Тэд төмс, амтат төмс, мандарваа цэцэг гэх мэт ургадаг.
агаарын үндэс
Эдгээр нь агаарын хэсэгт ургадаг хажуугийн үндэс юм. Тоогоор илэрхийлнэ халуун орны ургамал. Ус, хүчилтөрөгчийг агаараас шингээж авдаг. Дутмаг нөхцөлд ургадаг халуун орны ургамалд байдаг ашигт малтмал.

амьсгалын замын үндэс
Энэ нь субстрат, усны гадаргуугаас дээш өргөгдөж, дээшээ ургадаг нэг төрлийн хажуугийн үндэс юм. Ийм төрлийн үндэс нь хэт чийглэг хөрсөн дээр, намаг нөхцөлд ургадаг ургамалд үүсдэг. Ийм үндэсийн тусламжтайгаар ургамал нь агаараас дутагдаж буй хүчилтөрөгчийг хүлээн авдаг.
Дэмжих (самбар хэлбэртэй) үндэс
Энэ төрлийн модны үндэс нь том зүйлийн (гаш, хайлаас, улиас, халуун орны гэх мэт) онцлог шинж чанартай бөгөөд тэдгээр нь хажуугийн үндэсээр үүссэн, хөрсний гадаргуугийн ойролцоо буюу түүнээс дээш дамждаг гурвалжин босоо ургамлууд юм. Тэдгээр нь модонд тулгуурласан самбартай төстэй тул тэдгээрийг самбар хэлбэртэй гэж нэрлэдэг.
Сорох үндэс (хаусториа)
Энэ бол авирах ургамлын иш дээр үүсдэг нэмэлт нэмэлт үндэс юм. Тэдгээрийн тусламжтайгаар ургамал нь тодорхой тулгуурт бэхлэх, авирах (нэхэх) чадвартай байдаг. Ийм үндэс нь жишээлбэл, бат бөх ficus, ivy гэх мэт байдаг.

Эвхэгддэг (агшилт) үндэс
Ургамлын шинж чанар, үндэс нь суурь дээр уртааш чиглэлд огцом буурдаг. Жишээ нь булцуутай ургамал байж болно. Эвхэгддэг үндэс нь булцуу, эх үр тариаг хөрсөн дэх зарим завсарлагатай болгодог. Нэмж дурдахад, тэдгээрийн оршихуй нь сарнай цэцэг (жишээлбэл, данделион) газарт нягт таарч, босоо үндэслэг иш, эх хүзүүвчний газар доорх байрлалыг тодорхойлдог.
Микориза (мөөгөнцрийн үндэс)
Микориза нь мөөгөнцрийн гифтэй дээд ургамлын үндэсийг сүлжиж, үндэс үсний үүрэг гүйцэтгэдэг симбиоз (харилцан ашигтай хамтын амьдрал) юм. Мөөгөнцөр нь ургамлыг ус, түүнд ууссан шим тэжээлээр хангадаг. Ургамал нь эргээд мөөгөнцөрийг амин чухал үйл ажиллагаанд нь шаардлагатай органик бодисоор хангадаг.
Микориза нь олон өндөр ургамал, ялангуяа модлог ургамлын үндэст байдаг.
бактерийн зангилаа
Эдгээр нь азотыг тогтоогч бактеритай симбиотик амьдрахад тохирсон өөрчилсөн хажуугийн үндэс юм. Зангилаа үүсэх нь залуу үндсийг дотоод засал руу нэвчсэний улмаас үүсдэг. Ийм харилцан ашигтай хамтын амьдрал нь ургамалд азотыг хүлээн авах боломжийг олгодог бөгөөд үүнийг бактери агаараас тэдэнд хүртээмжтэй хэлбэрт шилжүүлдэг. Харин нянгууд нь бусад төрлийн бактеритай өрсөлдөхгүйгээр ажиллах боломжтой тусгай амьдрах орчинтой байдаг. Үүнээс гадна тэд ургамлын үндэст агуулагдах бодисыг ашигладаг.

Бактерийн зангилаа нь хөрсийг азотоор баяжуулахын тулд тариалангийн эргэлтэнд нөхөн сэргээх бодис болгон өргөн ашигладаг буурцагт ургамлын овгийн ургамлуудын хувьд ердийн зүйл юм. Хөх, шар царгас, улаан ба эспарцет, эвэрт царцаа гэх мэт үндэстний буурцагт ургамлууд нь азотыг хамгийн сайн шингээгч ургамалд тооцогддог.
Дээрх хувиралтаас гадна тулгуур үндэс (ишийг бэхжүүлэхэд тусалдаг), ганзагатай үндэс (ургамлыг шингэн шаварт живүүлэхгүй байх), үндэс сорогч (нахиа нахиатай, ургамлын үржлийг хангадаг) зэрэг өөр төрлийн үндэс байдаг.
Филогенетикийн хувьд үндэс нь иш, навчнаас хожуу үүссэн - ургамал газар дээрх амьдралд шилжсэнтэй холбогдуулан үндэстэй газар доорх мөчрүүдээс үүссэн байж магадгүй юм. Үндэс нь тодорхой дарааллаар байрлуулсан навч, нахиагүй байдаг. Энэ нь оройн ургалтаар тодорхойлогддог бөгөөд түүний хажуугийн мөчрүүд нь дотоод эдээс үүсдэг, өсөлтийн цэг нь эх малгайгаар хучигдсан байдаг. Үндэс систем нь ургамлын организмын амьдралын туршид үүсдэг. Заримдаа үндэс нь шим тэжээлийн хангамжид хуримтлагдах газар болж чаддаг. Энэ тохиолдолд үүнийг өөрчилдөг.
Үндэс төрлүүд
Үндсэн үндэс нь үрийн соёололтын үед үр хөврөлийн үндэсээс үүсдэг. Энэ нь хажуугийн үндэстэй.
Адал үндэс нь иш, навч дээр үүсдэг.
Хажуугийн үндэс нь аливаа үндэсийн салбар юм.
Үндэс бүр (үндсэн, хажуугийн, гэнэтийн) салбарлах чадвартай бөгөөд энэ нь үндэс системийн гадаргууг ихээхэн хэмжээгээр нэмэгдүүлдэг бөгөөд энэ нь хөрсөн дэх ургамлыг илүү сайн бэхжүүлж, тэжээллэг чанарыг нь сайжруулдаг.
Үндэс системийн төрлүүд
Үндэс систем нь сайн хөгжсөн үндсэн үндэстэй үндэс ба утаслаг гэсэн хоёр үндсэн төрөл байдаг. Шилэн үндэс систем нь ижил хэмжээтэй олон тооны гэнэтийн үндэсээс бүрддэг. Үндэсний бүх масс нь хажуугийн эсвэл гэнэтийн үндэсээс бүрдэх ба дэлбээ шиг харагддаг.
Өндөр салаалсан үндэс систем нь асар том шингээгч гадаргууг бүрдүүлдэг. Жишээлбэл,
- өвлийн хөх тарианы үндэс нийт урт 600 км хүрдэг;
- үндэс үсний урт - 10,000 км;
- үндэсийн нийт гадаргуу нь 200 м 2 байна.
Энэ нь газрын дээрх массын талбайгаас хэд дахин их юм.

Хэрэв ургамал нь сайн тодорхойлогдсон үндсэн үндэстэй бөгөөд гэнэтийн үндэс үүсвэл холимог төрлийн үндэс систем (байцаа, улаан лооль) үүсдэг.
Үндэсний гадаад бүтэц. Үндэсний дотоод бүтэц
Үндэс бүсүүд

үндэс таг
Үндэс нь боловсролын эдийн залуу эсүүд байрладаг үзүүрээрээ урт ургадаг. Өсөн нэмэгдэж буй хэсэг нь үндэсийн үзүүрийг гэмтээхээс хамгаалж, ургах явцад хөрсөнд үндэсийн хөдөлгөөнийг хөнгөвчлөх үндэсний малгайгаар хучигдсан байдаг. Сүүлчийн функц нь үндэс ба хөрсний хэсгүүдийн хоорондох үрэлтийг багасгадаг салст бүрхэвчээр бүрхэгдсэн эх малгайны гадна талын хананы өмчийн улмаас хийгддэг. Тэд бүр хөрсний тоосонцорыг түлхэж чаддаг. Үндэс малгайны эсүүд нь ихэвчлэн цардуулын үр тариа агуулсан амьд байдаг. Малгайны эсүүд хуваагдсаны улмаас байнга шинэчлэгддэг. Эерэг геотропик урвалд оролцдог (үндэс ургах дэлхийн төв рүү чиглэсэн чиглэл).

Хуваалтын бүсийн эсүүд идэвхтэй хуваагддаг бөгөөд энэ бүсийн урт нь өөр өөр зүйл, нэг ургамлын өөр өөр үндэст өөр өөр байдаг.
Хуваах бүсийн ард өргөтгөлийн бүс (өсөлтийн бүс) байдаг. Энэ бүсийн урт нь хэдэн миллиметрээс хэтрэхгүй.
Шугаман өсөлт дуусахад үндэс үүсэх гурав дахь үе шат эхэлдэг - түүний ялгаралт, эсийн ялгарах, мэргэшсэн бүс (эсвэл үндэс үс, шингээлтийн бүс) үүсдэг. Энэ бүсэд эпиблемийн гаднах давхарга (rhizoderm) үндэс үстэй, анхдагч бор гадаргын давхарга, төв цилиндр аль хэдийн ялгагдана.
Үсний үндэсийн бүтэц
Үндэс үс нь үндсийг бүрхсэн гаднах эсийн өндөр сунасан ургалт юм. Үндэс үсний тоо маш их (1 мм2 талбайд 200-300 үс). Тэдний урт нь 10 мм хүрдэг. Үс маш хурдан үүсдэг (алимны модны залуу суулгацанд 30-40 цагийн дотор). Үндэс үс нь богино насалдаг. Тэд 10-20 хоногийн дотор үхэж, шинэ үндэс нь залуу хэсэгт ургадаг. Энэ нь хөрсний шинэ давхрага үүсэхийг үндэс болгон баталгаажуулдаг. Үндэс нь тасралтгүй ургаж, үндэс үсний илүү олон шинэ хэсгүүдийг үүсгэдэг. Үс нь зөвхөн бодисын бэлэн уусмалыг шингээж авахаас гадна хөрсний тодорхой бодисыг уусгахад хувь нэмрээ оруулж, дараа нь шингээж чаддаг. Үндэсний үс унжсан хэсэг хэсэг хугацаанд ус шингээх чадвартай боловч дараа нь үйсэн бүрхэж, энэ чадвараа алддаг.
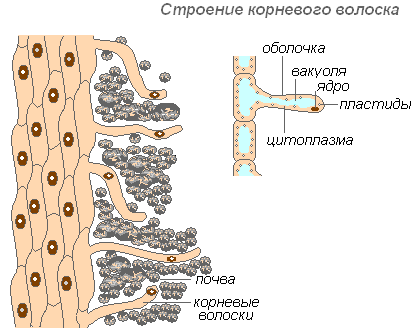
Үсний бүрээс нь маш нимгэн тул шим тэжээлийг шингээхэд тусалдаг. Бараг бүх үсний эсийг цитоплазмын нимгэн давхаргаар хүрээлэгдсэн вакуоль эзэлдэг. Цөм нь эсийн дээд хэсэгт байрладаг. Эсийн эргэн тойронд салст бүрхэвч үүсдэг бөгөөд энэ нь үндэс үсийг хөрсний тоосонцортой нааж, тэдгээрийн холбоог сайжруулж, системийн гидрофилийг нэмэгдүүлдэг. Шингээлт нь эрдэс давсыг уусгадаг үндэс үсээр хүчил (нүүрстөрөгч, алим, нимбэг) ялгарах замаар хөнгөвчилдөг.
Үндэс үс нь механик үүрэг гүйцэтгэдэг - тэдгээр нь хөрсний хэсгүүдийн хооронд дамждаг үндэсийн дээд хэсэгт дэмжлэг болдог.
Микроскопоор шингээх бүс дэх үндэсийн хөндлөн огтлол дээр түүний бүтэц нь эсийн болон эдийн түвшин. Үндэсний гадаргуу дээр үндэслэг иш, доор нь холтос байдаг. Cortex-ийн гаднах давхарга нь экзодерм, дотроос гол паренхим юм. Түүний нимгэн ханатай амьд эсүүд нь хадгалах функцийг гүйцэтгэдэг, шим тэжээлийн уусмалыг радиаль чиглэлд - шингээгч эдээс модны судас хүртэл явуулдаг. Тэд мөн ургамалд шаардлагатай олон тооны органик бодисуудыг нэгтгэдэг. Cortex-ийн дотоод давхарга нь эндодерм юм. Эндодермийн эсүүдээр дамжин бор гадаргаас төв цилиндрт орж ирдэг шим тэжээлийн уусмалууд нь зөвхөн эсийн протопластаар дамждаг.

Холтос нь үндэсийн төв цилиндрийг хүрээлдэг. Энэ нь удаан хугацааны туршид хуваагдах чадварыг хадгалдаг эсийн давхаргатай хиллэдэг. Энэ бол перицикл юм. Перицикл эсүүд нь хажуугийн үндэс, adnexal нахиа, дунд боловсролын эд эсийг үүсгэдэг. Перициклээс дотогшоо, үндэсний төвд дамжуулагч эдүүд байдаг: баст ба мод. Тэд хамтдаа радиаль дамжуулагч цацраг үүсгэдэг.
Үндэс дамжуулах систем нь ус, эрдэс бодисыг үндэснээс иш рүү (дээш гүйдэл), органик бодисыг ишнээс үндэс рүү (доошоо урсгал) дамжуулдаг. Энэ нь судасны фиброз багцуудаас бүрдэнэ. Багцын гол бүрэлдэхүүн хэсэг нь флоемын хэсгүүд (үндсээр дамжин бодисууд үндэс рүү шилждэг) ба ксилем (үндэсээс бодисууд шилждэг) юм. Флоемын гол дамжуулагч элементүүд нь шигшүүр хоолой, ксилем нь гуурсан хоолой (судас) ба трахеид юм.
Үндсэн амьдралын үйл явц
Усны тээвэрлэлтийг үндэс
Хөрсний шим тэжээлийн уусмалаас үндсийн үсээр ус шингээж, эндодермис дэх дамжих эсүүдээр дамжин радиаль судасны багцын ксилем рүү анхдагч бор гадаргын эсийн дагуу радиаль чиглэлд дамждаг. Үсний үсээр ус шингээх эрчмийг сорох хүч (S) гэж нэрлэдэг бөгөөд энэ нь осмотик (P) ба тургор (T) даралтын зөрүүтэй тэнцүү байна: S=P-T.
Осмосын даралт нь тургорын даралттай (P=T) тэнцүү байх үед S=0 байвал үндэс үсний эс рүү ус урсахаа болино. Хэрэв хөрсний шим тэжээлийн уусмал дахь бодисын агууламж эсийн доторхоос өндөр байвал ус эсийг орхиж, плазмолиз үүсэх болно - ургамал хатах болно. Энэ үзэгдэл нь хуурай хөрсний нөхцөлд, түүнчлэн ашигт малтмалын бордоог хэтрүүлэн хэрэглэх үед ажиглагддаг. Үндэс эс дотор үндэсийн сорох хүч нь үндэслэг давхаргаас төв цилиндр рүү нэмэгддэг тул ус нь концентрацийн градиент дагуу (өөрөөр хэлбэл өндөр концентрацитай газраас бага концентрацитай газар руу) шилжиж, үндэс даралтыг үүсгэдэг. Энэ нь ксилемийн судаснуудын дагуу усны баганыг босгож, дээш чиглэсэн урсгал үүсгэдэг. Энэ нь "шүүс" хурааж авах үед хаврын навчгүй их бие, эсвэл зүссэн хожуул дээр олддог. Мод, шинэ хожуул, навчнаас усны урсацыг ургамлын "уйлах" гэж нэрлэдэг. Навч цэцэглэх үед тэд мөн сорох хүчийг бий болгож, усыг өөртөө татдаг - хөлөг онгоц бүрт тасралтгүй усны багана үүсдэг - хялгасан судасны хурцадмал байдал. Үндэсний даралт нь усны урсгалын доод хөдөлгүүр, навчны сорох хүч нь дээд хэсэг юм. Энгийн туршилтын тусламжтайгаар та үүнийг баталж чадна.
Үндэс нь ус шингээх
Зорилтот:язгуурын үндсэн үүргийг олж мэд.
Бидний хийдэг зүйл:нойтон модны үртэс дээр ургасан ургамал, түүний үндэс системийг сэгсэрч, үндсийг нь аяга ус руу буулгана. Ууршилтаас хамгаалахын тулд усан дээр нимгэн давхарга хийнэ. ургамлын тосмөн түвшинг тэмдэглэ.

Бидний ажиглаж буй зүйл:нэг эсвэл хоёр хоногийн дараа савны ус тэмдэгтээс доош унав.

Үр дүн:Тиймээс үндэс нь усыг сорж, навч руу авчирдаг.
Үндэс нь шим тэжээлийг шингээж авдаг болохыг нотлох өөр нэг туршилтыг хийж болно.
Бидний хийдэг зүйл:бид ургамлын ишийг тайрч, 2-3 см өндөр хожуул үлдээж, хожуул дээр 3 см урт резинэн хоолой тавьж, дээд үзүүрт нь 20-25 см өндөр муруй шилэн хоолой тавина.
Бидний ажиглаж буй зүйл:шилэн хоолой дахь ус дээшээ гарч урсдаг.
Үр дүн:Энэ нь үндэс нь хөрсний усыг ишэнд шингээж авдгийг нотолж байна.

Усны температур нь үндсээр нь ус шингээх хурдад нөлөөлдөг үү?
Зорилтот:Температур нь үндэсийн үйл ажиллагаанд хэрхэн нөлөөлдөг болохыг олж мэдээрэй.
Бидний хийдэг зүйл:нэг шил бүлээн усаар (+17-18ºС), нөгөө нь хүйтэн усаар (+1-2ºС) байх ёстой.
Бидний ажиглаж буй зүйл:Эхний тохиолдолд ус их хэмжээгээр ялгардаг, хоёрдугаарт - бага зэрэг эсвэл бүрмөсөн зогсдог.
Үр дүн:Энэ нь температур нь үндэсийн гүйцэтгэлд хүчтэй нөлөөлдөг гэдгийг нотолж байна.

Халуун ус нь үндэсээр идэвхтэй шингэдэг. Үндэсний даралт нэмэгддэг.

Хүйтэн ус нь үндэст муу шингэдэг. Энэ тохиолдолд үндэс даралт буурдаг.
эрдэс тэжээл
Ашигт малтмалын физиологийн үүрэг маш их. Эдгээр нь синтезийн үндэс суурь болдог органик нэгдлүүд, түүнчлэн коллоидуудын физик төлөвийг өөрчилдөг хүчин зүйлүүд, i.e. протопластын бодисын солилцоо, бүтцэд шууд нөлөөлдөг; биохимийн урвалын катализаторын үүрэг гүйцэтгэдэг; эсийн тургор, протоплазмын нэвчилтэд нөлөөлөх; ургамлын организм дахь цахилгаан ба цацраг идэвхт үзэгдлийн төвүүд юм.
Шим тэжээлийн уусмалд азот, фосфор, хүхэр, дөрвөн металл - кали, магни, кальци, төмөр зэрэг гурван металл бус бодис байгаа тохиолдолд л ургамлын хэвийн хөгжил боломжтой болох нь тогтоогдсон. Эдгээр элемент бүр өөрийн гэсэн утгатай бөгөөд өөр зүйлээр сольж болохгүй. Эдгээр нь макронутриентууд бөгөөд тэдгээрийн ургамал дахь концентраци 10 -2 -10% байдаг. Ургамлын хэвийн хөгжилд микроэлементүүд шаардлагатай байдаг бөгөөд тэдгээрийн концентраци нь эсэд 10 -5 -10 -3% байдаг. Эдгээр нь бор, кобальт, зэс, цайр, манган, молибден гэх мэт Эдгээр бүх элементүүд нь хөрсөнд байдаг боловч заримдаа хангалтгүй хэмжээгээр байдаг. Тиймээс хөрсөнд эрдэс болон органик бордоог хэрэглэдэг.
Үндэс хүрээлэн буй орчин нь шаардлагатай бүх шим тэжээлийг агуулсан байвал ургамал хэвийн ургаж, хөгждөг. Хөрс бол ихэнх ургамлын ийм орчин юм.
Үндэс амьсгал
Ургамлын хэвийн өсөлт, хөгжилд цэвэр агаар нь үндэс рүү орох шаардлагатай. Тийм эсэхийг шалгацгаая?
Зорилтот:үндэс агаар хэрэгтэй юу?
Бидний хийдэг зүйл:Устай хоёр ижил савыг авцгаая. Бид хөгжиж буй суулгацыг хөлөг онгоц бүрт байрлуулна. Бид өдөр бүр нэг савны усыг шүршигч сав ашиглан агаараар дүүргэдэг. Хоёр дахь саванд байгаа усны гадаргуу дээр ус руу орох агаарын урсгалыг хойшлуулдаг тул ургамлын тосыг нимгэн давхаргаар хийнэ.

Бидний ажиглаж буй зүйл:хэсэг хугацааны дараа хоёр дахь саванд байгаа ургамал ургахаа больж, хатаж, эцэст нь үхэх болно.

Үр дүн:ургамлын үхэл нь үндэс амьсгалахад шаардлагатай агаар дутагдсанаас болдог.
Үндэс өөрчлөлтүүд
Зарим ургамалд нөөц тэжээл нь үндэст хуримтлагддаг. Тэд нүүрс ус, эрдэс давс, витамин болон бусад бодисыг хуримтлуулдаг. Ийм үндэс нь зузаан нь хүчтэй ургадаг бөгөөд ер бусын дүр төрхийг олж авдаг. Үндэс ба иш нь үндэс үр тариа үүсгэхэд оролцдог.
Үндэс
Хэрэв нөөцийн бодисууд үндсэн үндэс болон үндсэн найлзууруудын ишний ёроолд хуримтлагдвал эх үр тариа (лууван) үүсдэг. Үндэс үүсгэгч ургамал нь ихэвчлэн хоёр наст ургамал юм. Амьдралын эхний жилд тэд цэцэглэдэггүй, эх үр тарианд маш их шим тэжээлийг хуримтлуулдаг. Хоёр дахь нь тэд хуримтлагдсан шим тэжээлийг ашиглан хурдан цэцэглэж, жимс жимсгэнэ, үрийг үүсгэдэг.
үндэс булцуу

Dahlia-д нөөц бодисууд гэнэтийн үндэст хуримтлагдаж, үндэс булцууг үүсгэдэг.
бактерийн зангилаа
Гэрийн хошоонгор, люпин, царгасны хажуугийн үндэс нь өвөрмөц өөрчлөгддөг. Бактери нь залуу хажуугийн үндэст суурьшдаг бөгөөд энэ нь хөрсний агаараас хийн азотыг шингээхэд хувь нэмэр оруулдаг. Ийм үндэс нь зангилаа хэлбэртэй байдаг. Эдгээр бактерийн ачаар эдгээр ургамлууд азотын дутагдалтай хөрсөнд амьдарч, илүү үржил шимтэй болгодог.
ганган

Далайн завсрын бүсэд ургадаг налуу нь ганган үндэс үүсгэдэг. Усан дээгүүр өндөр, тогтворгүй шаварлаг газар том навчит найлзуурыг барьдаг.
Агаар

Модны мөчир дээр амьдардаг халуун орны ургамал нь агаарын үндсийг хөгжүүлдэг. Тэд ихэвчлэн цахирмаа, бромелиад, зарим оймын модноос олддог. Агаарын үндэс нь агаарт чөлөөтэй унжиж, газарт хүрч, бороо, шүүдэр унасан чийгийг шингээдэггүй.
Ретраторууд

Булцуут болон хясааны ургамалд, жишээлбэл, матарт, олон тооны утас шиг үндэстэй, хэд хэдэн зузаан, ухрах үндэс гэж нэрлэгддэг. Бууруулах нь ийм үндэс нь corm-ыг хөрсөнд илүү гүнзгийрүүлдэг.
Багана хэлбэртэй

Фикус нь газрын дээрх булчирхайлаг үндэс буюу тулгуур үндэсийг хөгжүүлдэг.
Үндэс амьдрах орчин болох хөрс
Ургамлын хөрс нь ус, шим тэжээлийг хүлээн авдаг орчин юм. Хөрсөн дэх эрдэс бодисын хэмжээ нь эх чулуулгийн өвөрмөц онцлог, организмын идэвхжил, ургамлын өөрийнх нь амин чухал үйл ажиллагаа, хөрсний төрлөөс хамаарна.
Хөрсний тоосонцор чийгийн төлөө үндэстэй өрсөлдөж, гадаргуу дээрээ хадгалдаг. Энэ нь гигроскопийн болон хальсанд хуваагддаг холбосон ус гэж нэрлэгддэг ус юм. Үүнийг молекулын таталцлын хүчээр барьдаг. Ургамлын чийгийг капилляр усаар төлөөлдөг бөгөөд энэ нь хөрсний жижиг нүхэнд төвлөрдөг.
Хөрсний чийг ба агаарын фазын хооронд антагонист харилцаа үүсдэг. Хөрсөнд том нүхтэй байх тусам эдгээр хөрсний хийн горим сайн байх тусам хөрсний чийг бага хадгалагдана. Ус ба агаар нь нэгэн зэрэг байрладаг, бие биедээ саад болохгүй бүтцийн хөрсөнд ус-агаарын хамгийн таатай горимыг хадгалдаг - ус нь бүтцийн дүүргэгч доторх хялгасан судсыг дүүргэж, тэдгээрийн хоорондох том нүхийг агаараар дүүргэдэг.
Ургамал ба хөрсний харилцан үйлчлэлийн шинж чанар нь хөрсний шингээх чадвар - химийн нэгдлүүдийг хадгалах эсвэл холбох чадвараас ихээхэн хамаардаг.
Хөрсний микрофлор нь органик бодисыг илүү ихээр задалдаг энгийн холболтууд, хөрсний бүтэц үүсэхэд оролцдог. Эдгээр үйл явцын шинж чанар нь хөрсний төрөл, ургамлын үлдэгдэл химийн найрлага, бичил биетний физиологийн шинж чанар болон бусад хүчин зүйлээс хамаарна. Хөрсний амьтад хөрсний бүтцийг бүрдүүлэхэд оролцдог: анелид, шавьжны авгалдай гэх мэт.
Биологийн болон хосолсон үр дүнд химийн процессуудхөрсөнд "ялзмаг" гэсэн нэр томъёогоор нийлдэг органик бодисын цогц цогцолбор үүсдэг.
Усны соёлын арга
Ургамалд ямар давс хэрэгтэй, түүний өсөлт хөгжилтөд ямар нөлөө үзүүлдэг вэ гэдгийг усны өсгөвөрт хийсэн туршилтаар тогтоосон. Усны соёлын арга нь ургамлыг хөрсөнд биш, харин эрдэс давсны усан уусмалд тариалах явдал юм. Туршилтын зорилгоос хамааран та уусмалаас тусдаа давсыг хасах, түүний агууламжийг багасгах эсвэл нэмэгдүүлэх боломжтой. Азот агуулсан бордоо нь ургамлын өсөлтийг, фосфор агуулсан бордоо нь жимс жимсгэний хамгийн эрт боловсорч гүйцээд, кали агуулсан бордоо нь навчнаас үндэс хүртэл органик бодисыг хамгийн хурдан гадагшлуулдаг болохыг тогтоожээ. Үүнтэй холбогдуулан азот агуулсан бордоог тарихаас өмнө эсвэл зуны эхний хагаст, фосфор, кали агуулсан бордоог зуны хоёрдугаар хагаст хэрэглэхийг зөвлөж байна.
Усны өсгөвөрлөх аргыг ашиглан ургамлын макро элементүүдийн хэрэгцээг тогтоох төдийгүй янз бүрийн микроэлементүүдийн үүргийг олж мэдэх боломжтой байв.
Одоогийн байдлаар ургамлыг гидропоник, аэропоник аргаар ургуулах тохиолдол байдаг.
Гидропоник бол хайрга дүүргэсэн саванд ургамал тариалах явдал юм. Шим тэжээлийн уусмал агуулсан шаардлагатай элементүүд, доороос нь савнууд руу тэжээгддэг.
Аэропоник бол ургамлын агаарын соёл юм. Энэ аргын тусламжтайгаар эх систем нь агаарт байгаа бөгөөд автоматаар (нэг цагийн дотор хэд хэдэн удаа) шим тэжээлийн давсны сул уусмалаар шүршинэ.
ОРГАНографи
Орган зүй - амьд организмын эрхтнүүдийн тодорхойлолт.
Эрхтэн- морфологийн болон функциональ байдлаараа ялгаатай биеийн хэсэг. Тухайн ургамлын амьдралыг хадгалах, дараагийн хэд хэдэн үед нөхөн үржих гэсэн хоёр үндсэн чиг үүргийн дагуу ургамлын болон нөхөн үржихүйн эрхтнүүдийг ялгадаг.
Нэгдүгээрт, бид найлзуурууд болон үндэс системийг бүрдүүлдэг ургамлын эрхтнүүдийг судлах болно. Үндэс системийн элемент нь үндэс юм.
Үндэс
Үндэс нь найлзуураас хожуу хөгжиж, хуурай газрын анхны ургамал болох ринофитуудын үндэслэг ишнээс гаралтай.
Үндэсний өөр өөр хэсгүүд нь өөр өөр функцийг гүйцэтгэдэг бөгөөд тодорхой шинж чанартай байдаг морфологийн онцлог. Эдгээр хэсгүүдийг бүс гэж нэрлэдэг. Залуу үндэсийн бүсүүд: хуваагдах бүс, өсөлт эсвэл сунах бүс, шингээх эсвэл шингээх бүс, дамжуулах бүс.
Гадна оройн меристем нь эх малгайгаар бүрхэгдсэн байдаг. Малгайны амьд паренхим эсүүд салиа үүсгэдэг. Салст бодис нь өндөр чийгшилтэй пектин полисахарид юм. Энэ нь диктиосомын цэврүүтүүдэд хуримтлагддаг бөгөөд энэ нь плазмын мембрантай нийлж, плазмын мембран ба эсийн мембран хоорондын зайд ялгардаг. Дараа нь салиа нь эсийн мембраны гадаргуу дээр гарч, жижиг дусал үүсгэдэг.
Энэхүү салст бүрхүүлийн улмаас хөрсний тоосонцор нь үндэсийн үзүүр, үсний үсэнд наалддаг. Салс нь ион солилцооны шингээгч гадаргуу болж үйлчилдэг. Салст бүрхэвч нь үндэсийн үзүүрийг хамгаалдаг хортой бодисуудхөрс, хуурайшилтаас болж ашигтай нян суурьших таатай нөхцлийг бүрдүүлдэг.
Үндэс ургаж, малгайгаа урагш түлхэхэд малгайны захын эсүүд ховхордог. Харагдахаас эхлээд гуужих хүртэлх эд эсийн амьдрах хугацаа нь малгайны урт, ургамлын төрлөөс хамаарч 4-9 хоног байна.
Root cap нь эх газрын хүлээн авах ажиллагааг хянадаг. Малгай дахь хүндийн хүчийг мэдрэх газар нь эсийн төв багана - колумелла бөгөөд амилопластууд нь статолит (таталцлын рецептор) үүрэг гүйцэтгэдэг.
Оройн меристем нь нягт цитоплазм, том цөм бүхий жижиг (10-20 мкм диаметртэй) олон талт эсүүдээс тогтдог.
Спорын ургамал нь нэг төрлийн үндэс оройтой байдаг.
Үрийн ургамлын үндэст оройн хоёр үндсэн зохион байгуулалтыг тодорхойлсон. Тэдгээрийн аль нэгэнд нь бүх хэсэгт нийтлэг эхний үсэг байдаг. Энэ нь оройн меристемийн нээлттэй төрөл бөгөөд филогенетикийн хувьд анхдагч байж магадгүй юм. Хоёр дахь төрөлд гурван хэсэг тус бүр, i.e. төв (эсвэл дамжуулагч) цилиндр, анхдагч холтос ба үндэсийн таг нь өөрийн гэсэн эхний үсэгтэй. Энэ бол оройн меристемийн хаалттай төрөл юм.
Эхний үе шатанд эсийн өсөлтийг зохион байгуулах үүрэгтэй эхний үсэг нь үндэс өсөлтийн дараагийн үе шатанд митозын идэвхээ алддаг. Үүний оронд эх биеийн гүнд байрлах эсүүд митозын үйл ажиллагааг харуулдаг. Эдгээр ажиглалтууд нь оройн меристем дэх амралтын төвийн онолыг бий болгосон. Энэ онолын дагуу үндэс бие дэх хамгийн алслагдсан эсүүд (төв цэгээс хамгийн алслагдсан эрхтний алслагдсан хэсэг: язгуурын доод хэсэг) (плерома ба периблемагийн эхний үсэг) хуваагдах нь ховор, ойролцоогоор ижил байдаг. хэмжээтэй, NA болон уургийн нийлэгжилт багатай байдаг. Амрах төв нь хагас бөмбөрцөг эсвэл диск хэлбэртэй, үндэсийн хэмжээнээс хамаарч өөр өөр хэмжээтэй байдаг. Амрах төвийн эсүүд хөрш зэргэлдээх эсүүдээс ойролцоогоор 10 дахин удаан хуваагддаг.
Туршилтаар гэмтсэн үндэст унтаа төв нь меристемийг сэргээх чадвартай. Өөрөөр хэлбэл, амрах төв нь бүтцийг бий болгох, үндсийг хөгжүүлэхэд чухал үүрэг гүйцэтгэдэг.
Үндэс үзүүрийн онцлог нь оройн меристемээс үүссэн уртааш эгнээний эсүүдийг байрлуулах явдал юм.
Үндэсний оройн меристемийн давхаргууд:
Дерматоген(Грекээр derma - арьс, genos - төрөлт) - оройн меристемийн гадна давхарга нь үндэслэг давхаргад хуваагдана. Зөв хэлбэрийн дерматоген эсүүд.
Плерома(Грек плерома - дүүргэх) - оройн меристемийн төв хэсэг нь төв цилиндрийн эсүүдэд хуваагддаг. Эсүүд тэнхлэгийн дагуу сунадаг.
аюул(Грек periblema - бүрхэвч) - дерматоген ба плеромын хоорондох оройн меристемийн давхарга нь анхдагч бор гадаргын эсүүдэд хуваагддаг.
Анхан шатны кортекс нь суналтын бүсийн түвшинд ялгаатай байдаг. Колоничлолын бүсэд анхдагч кортекс нь хөндлөн огтлолын ихэнх хэсгийг эзэлдэг. Анхдагч меристемээс гаралтай тул "анхдагч", захын байрлалтай тул "холтос" гэж нэрлэдэг.
Rhizoderm доор байрлах анхдагч бор гадаргын гаднах эсийг экзодерм гэж нэрлэдэг. Анхдагч бор гадаргын дотоод давхарга нь эндодерм юм.
Анхдагч бор гадаргын ихэнх хэсэг нь мезодерм юм. Мезодерм нь сул байрлалтай паренхим эсүүдээс бүрддэг бөгөөд энэ нь үндэс эсийг агааржуулахад хувь нэмэр оруулдаг.
Гимносперм ба ихэнх дикотуудын үндэс нь анхдагч холтосоо эрт хаядаг. Тэдгээрийн дотор анхдагч бор гадаргын эсүүд паренхим хэвээр үлддэг. Үүний эсрэгээр, монокотын анхдагч бор гадар нь үндэсийн амьдралын туршид хадгалагддаг бөгөөд түүний эсүүд нь хоёрдогч хүрэн бүрхүүл үүсгэдэг.
Үндэсний төвийг тэнхлэгийн цилиндр - стела эзэлдэг. Өсөлтийн бүсэд ялгарч эхэлдэг. Тэнхлэгийн цилиндр үүсэх нь түүний гаднах давхарга үүсэхээс эхэлдэг. периcycle.Перицикл нь анхдагч хажуугийн меристемд хамаардаг бөгөөд түүний эсүүд меристемийн үйл ажиллагааг удаан хугацаанд хадгалдаг. Ихэвчлэн перицикл нь нэг давхарга эсээр дүрслэгддэг боловч олон давхаргат байж болно.
Ихэнх үрийн ургамалд хажуугийн үндэс нь перицикл дотор тавигдаж, adnexal нахиа гарч ирдэг. Хоёрдогч өсөлттэй зүйлүүдийн хувьд перицикл нь камби үүсэхэд оролцдог бөгөөд феллогенийн эхний давхаргыг үүсгэдэг.
Перициклийн доор прокамбиал эсүүд тавигдаж, анхдагч флоем, дараа нь анхдагч ксилем үүсдэг. Янз бүрийн төрөл зүйлд ксилемийн туяаны тоо хоёр, гурав, дөрөвөөс хэд хэдэн хооронд хэлбэлздэг. Ксилемийн цацрагийн тоогоор үндсийг диарх, триарх, тетрарх, полиарх гэж нэрлэдэг. Тетрархын үндэс нь анхдагч гэж тооцогддог. Монокотууд нь полиарх стелетэй байдаг.
Тиймээс, ерөнхий төлөвлөгөөҮндэсний анхдагч бүтэц, эд эсийн морфологийн ялгаатай байдал нь үндэс нь ус, эрдэс бодисыг шингээх, дамжуулах үйл ажиллагаанд дасан зохицох өндөр түвшний мэргэшсэн байдлыг харуулж байна.
Хажуугийн үндэс үүсэх
Хажуугийн үндэс нь оройн меристемээс өөр өөр зайд төв цилиндрийн захад үүсдэг. Учир нь тэдгээр нь үндэсийн гүн байрлах эдэд байрладаг бөгөөд тэдгээрийн гарал үүслийг эндоген гэж нэрлэдэг (Грек хэлнээс "дотор төрсөн"). Гимносперм ба ангиоспермийн хажуугийн үндэс нь перицикл дотор тавигддаг. Хажуугийн үндэс үүсэхэд оролцдог перициклийн эсүүд нь үндэс нуман хаалга юм.
Мөн эндодермээс үндэс анхдагч эсийн хэд хэдэн давхаргыг үүсгэж болно.
Диарх үндэст флоем ба ксилемийн хооронд хажуугийн үндэс, ксилемийн эсрэг талд гурвалсан, тетрахник үндэс, монокотын полиарх үндэст флоемын эсрэг байрлана. Перицикл эсүүд өөр өөр чиглэлд хуваагддаг. Үүний үр дүнд жижиг үндэстэй булцуу үүсдэг. Хэмжээ ихсэх тусам үндэс нь эндодерм дээр дарагддаг. Гэхдээ эндодермийн бүрхүүлүүд хатуу байдаг. Тиймээс эндодерм нь цухуйсан байдаг. Эндодермийн товойсон хэсгийг үндэс халаас гэж нэрлэдэг.
Үндэсний анхдагч нь анхдагч бор гадаргаар дамжин өнгөрч, замын дагуух бор гадаргын эсийг устгадаг ферментүүдийг ялгаруулдаг. Хөгжлийн эхний үе шатанд энэ суурь нь эх малгай, оройн ба анхдагч меристемийг үүсгэдэг. Цаг хугацаа өнгөрөхөд паренхимийн эсүүд нь ксилем ба флоемын элементүүдэд хуваагддаг тул хажуугийн болон эх үндэсийн төв цилиндрүүд хоорондоо холбогддог.
Монокотуудад анхдагч үндэс бүтэц нь амьдралын туршид хадгалагдаж, хоёрдогч эдүүд үүсдэггүй. Нас ахих тусам эсийн хэмжээ нэмэгдэж, эсийн мембран өтгөрдөг боловч үндэс бүтцийн ерөнхий "төлөвлөгөө" өөрчлөгдөөгүй хэвээр байна.
Гимносперм ба дикотуудад хоёрдогч өөрчлөлт гарч, хоёрдогч үндэс бүтэц үүсдэг. Үндэс хоёрдогч бүтэц үүсэх нь юуны түрүүнд камбиумын үйл ажиллагаатай холбоотой байдаг. Камби нь анхдагч флоем ба анхдагч ксилемийн хооронд үл ялгагдах про-камбын эсүүдийн хуваагдалаар үүсдэг. Дараа нь ксилемийн цухуйсан хэсгүүдийн гадна байрлах перициклийн эсүүд нь мөн камбийн үйл ажиллагааг харуулдаг.
Камби нь хоёрдогч ксилем эсийг төвд, хоёрдогч флоем эсийг захад байрлуулдаг. Ийм байдлаар дамжуулагч багцууд үүсдэг. Тэд анхдагч ксилемийн туяатай адил үүсдэг. Өсөн нэмэгдэж буй хоёрдогч эдүүдийн даралтын дор анхдагч phloem-ийн эсүүд хавтгайрдаг. Анхдагч ксилем нь төв байрлалаа хадгалж, од шиг харагдана. Цаг хугацаа өнгөрөхөд анхдагч ксилем мөн устаж үгүй болдог.
Үндэсний хоёрдогч бүтцэд анхдагч эдүүд нь дамжуулагч элементийн үүрэг гүйцэтгэдэггүй. Хоёр чиглэлд бодисын гүйдэл нь хоёрдогч эдээр дамждаг.
Перициклээс үүссэн камби нь нээлттэй барьцааны багцуудын хооронд байрладаг өргөн паренхимийн туяа үүсгэдэг. Эдгээр нь үндсэн гол цацрагууд юм. Эдгээр нь үндэсийн төв хэсэг ба анхдагч бор гадаргын хоорондох физиологийн холболтыг хангадаг. Хожим нь хоёрдогч ксилем ба флоемыг холбосон нарийн хоёрдогч медуляр туяа үүсч болно.
Хоёрдогч өөрчлөлт нь зөвхөн төв цилиндрт тохиолддоггүй. Дамжуулагч эдүүдийн зузаан ихэссэнээс анхдагч бор гадаргын гадна тал руу түлхэж, эндодермтэй хамт урагдаж, урсдаг.
Перициклийн гадна хэсэгт феллоген үүсч, фелема гадагшаа, олон давхаргат фелодерм нь дотогшоо байрлана. Тиймээс хоёрдогч бүтэцтэй дикотуудын үндэс нь перидермээр бүрхэгдсэн байдаг.
Олон наст үндэст феллоген үндэс гүнд үүсдэг. Дараа нь үндэс нь ишний нэгэн адил ритидом үүсгэдэг.
Хоёрдогч үндэс ургах экологийн чухал талгэдэг нь нэг зүйлийн өөр өөр ургамлын үндэс байгалийн жамаар ургах чадвар юм. Үндэс нь хоорондоо шүргэлцдэг газар хоёрдогч өсөлтөөр нэгддэг. Үндэс хамтдаа ургах үед нийлсэн түншүүдийн хооронд дамжуулагч системийн тасралтгүй байдал тогтдог. Үндэс нийлсэний ачаар олон мод хоорондоо холбогддог бөгөөд хэрэв тэдгээрийн заримыг нь огтолж авбал хожуул нь удаан хугацаанд амьд үлддэг. Үндэсийг нэгтгэх нь халдварыг нэг модноос нөгөө мод руу хурдан дамжуулахад хувь нэмэр оруулдаг.
Үндэс систем дэх үндсийг ялгахянз бүрийн хэлбэрээр илэрдэг. Хамгийн чухал нь гол, хажуугийн болон нэмэлт үндэс гэж ялгах явдал юм.
Тусгай бүлэгт дээд спорын ургамлын оройн меристемд үүссэн үндсийг ялгадаг. Эдгээр нь хамгийн эртний язгуур бүлэг бөгөөд газар дээрх болон газар доорх тэнхлэгүүдийн нэг салаалсан системийн нэг хэсэг болох анхдагч үндэстэй хамгийн ижил төстэй байдлаа хадгалсан гэж үздэг. Энэ бүлэгт багтсан үндсийг апикоген гэж нэрлэдэг.
Хувьслын явцад үндэс систем үүсэх, томоохон системчилсэн бүлгүүдэд хамаарах дарааллаар бид гурван зүйлийг тоймлон гаргаж болно. үндэс системийн хувьслын түвшин.
I. Дээд спорын үндэс систем нь зөвхөн apigenic үндэс системээр төлөөлдөг бөгөөд бие биенээ дараалан орлуулдаг бөгөөд энэ нь үндэс системийг шинэчлэх үндсэн механизм юм. Салбарлах нь оройн эсвэл хажуу тал бөгөөд хоёр төрлийн салаа нь бие биенээсээ үл хамааран үүссэн байж магадгүй юм. Нарийн мэргэшилтэй метаморфоз, үндэс байхгүй.
P. Gymnosperms үндэс систем нь үр хөврөлийн үндэс нь үр, хоёр туйлт үр хөврөлийн харагдах холбоотой байж болох үндсэн үндэс байгаа нь ялгагдана. Шинэчлэлийн механизмд харьяа тушаалын хажуугийн үндсийг оруулаад зогсохгүй rhizogeneous adventitous үндэс (жишээ нь, үндэс дээрх нэмэлт үндэс, найлзуурууд дээрх гэнэтийн үндэс нь гимноспермүүдэд мэдэгдэхүйц хөгжлийг хүлээж аваагүй). Мэргэшсэн болон хувирсан үндэс нь ховор байдаг.
Ш. Ангиоспермийн үндэс систем нь өвөрмөц үндэс, мөн метаморфоз, нарийн мэргэшсэн үндэс хүчтэй хөгждөг онцлогтой. Шинэчлэх механизм нь олон янз бөгөөд үр дүнтэй байдаг.
Үндэс системийн метаморфозууд
Үндэс системийн метаморфоз гэдэг нь физиологийн үйл ажиллагааны өөрчлөлтөөс үүдэлтэй филогенетик болон удамшлын хувьд тогтворжсон анатомийн болон морфологийн шинж чанаруудын цогц юм. Метаморфоз (метаморфоз, Грек - хувиргалт) нь удамшлын түвшинд тохиолддог өөрчлөлтүүдийн үр дүн бөгөөд энэ нь эрхтэн, эрхтэн тогтолцоо, организмын бүхэлдээ морфологи, үйл ажиллагааны бүтцийн өөрчлөлтөд хүргэдэг.
I. Хэвийн бус функцүүдийн гүйцэтгэлтэй холбоотой метаморфозууд:
1) амьсгалын замын үндэс;
2) эпифитүүдийн агаарын үндэс;
3) ганган үндэс;
4) нөөц үндэс.
P. Хэт үйл ажиллагаатай холбоотой метаморфозууд:
1) үндэс үр тариа;
2) үндэс боргоцой;
3) үндэс зангилаа;
4) микориза.
Үндэс үр тариа нь хоёр наст хоёр наст ургамлын онцлог шинж юм. Энэ бол түр зуурын нэр юм. Тэд ургамлын эрхтнүүдээс үүсдэг бөгөөд жимс жимсгэнэтэй ямар ч холбоогүй байдаг. Ишний доод хэсэг ба үндэс нь үндэс үр тариа үүсгэхэд оролцдог.
Амьдралын эхний жилд ургамал ургадаг, үндэс ургац үүсдэг. Хоёр дахь жилдээ цэцэг, жимс жимсгэнэ, үр үүсдэг. Энэ нь эх үр тариа нь ургамлын унтаа үеийг шилжүүлж, дараа жил нь бүрэн хөгжүүлэх боломжийг олгодог бодисыг хангадаг эрхтэн юм гэсэн үг юм.
Үндэс төрлүүд:
1. Монокамбиал.
Анхны үндэс нь тэнхлэгийн цилиндр юм. Үндэс ургац үүсэх нь нэг камбийн цагиргийг эхлүүлэх, ажиллуулах, цаашдын паренхимизацитай холбоотой хоёрдогч өөрчлөлтөөс эхэлдэг.
Камби нь хоёрдогч ксилемийг дотогшоо, хоёрдогч флоемыг гадагш гаргадаг гэдгийг бид мэднэ. Анатомийн ялгаа нь хадгалалтын эд эсийн хөгжлийн газартай холбоотой байдаг. Хэрэв флоемд паренхимийн хүчтэй хөгжил үүсвэл флоемын төрлийн эх үр тариа үүсдэг - луувангийн төрөл. Хэрэв ксилемд - ксилемийн төрөл - манжингийн төрөл.
2.Үндэс үр тарианы поликамбын төрөл.
Нэг ердийн камбийн цагираг үүсдэг, үлдсэн хэсэг нь перициклик гаралтай нэмэлтүүд юм. Тэдгээрийн тоо 25 хүртэл байж болно.Эдгээр камбийн нэмэлт цагиргууд нь нээлттэй барьцааны судасны багцуудыг үүсгэдэг. Бөгж нь эхний камбын цагирагаас хол байх тусам илүү их хадгалалтын паренхим үүсдэг. Поликамбын үндэс ургацын жишээ бол манжин юм.
Моно болон поликамбын үндэст үр тарианы анхдагч холтос нь эрт үхэж, үрэвсдэг. Бүрхүүл нь перидерм юм.
Тиймээс бүх эх үр тарианы нийтлэг шинж чанар нь дамжуулагч эдээр нэвчсэн их хэмжээний хадгалалтын паренхим байдаг. Дамжуулагч ба хадгалалтын эдүүдийн хоорондын нягт холбоог хоёрдогч өсөлтийн янз бүрийн өөрчлөлтөөр хангадаг.
Зарим ургамалд хадгалалтын үндэс нь боргоцой хэлбэртэй хажуугийн болон нэмэлт үндэсээс үүсдэг. Монокотуудын (любка) үндэс боргоцой нь камбиал, анхдагч бүтэцтэй; dicots - cambial, хоёрдогч бүтэцтэй. Жишээ нь: флоэм төрлийн мандарваа цэцгийн үндэс боргоцой.
Үндэс боргоцой нь бүх гадаргуу дээр гэнэтийн үндэс үүсгэдэг ба дээд хэсэгт нахиа үүсдэг тул ургамлын нөхөн үржихүйн эрхтэн юм.
Ургамлын өсөлт нь ихэвчлэн тогтмол азотын хангалтгүй хангамжаар хязгаарлагддаг гэдгийг мэддэг. Тогтвортой N 2 молекул хэлбэрийн азот нь агаар мандлын 80% -ийг бүрдүүлдэг. Бэхэлгээ хийхээс өмнө (N 2-ыг аммиак NH 3 болгон бууруулах) энэ молекулыг хуваах шаардлагатай. Азотын бэхжилтийг хөрсний органик бодисыг хэрэглэдэг зарим чөлөөт амьд бактери гүйцэтгэдэг. Зарим бактери нь дээд ургамлын үндэс дээр суурьшдаг. Ургамал нь органик үндэсийн шүүрэл бүхий бактерийг "татдаг". Мөн бичил биетэн нь эргээд ургамлыг тогтмол азотоор хангадаг. Хоёр организмын энэхүү харилцан ашигтай холбоог симбиоз гэж нэрлэдэг. Ихэнх тохиолдолд үндэс нь Rhizobium төрлийн азотын нянтай холбоотой байдаг. Хоёр болон нэг наст ургамлын 60 гаруй гэр бүл нянтай симбиоз үүсэх боломжтой. Ургамлын төрөл бүр өөрийн гэсэн симбиотик Rhizobium-тай байдаг. Ургамал ба бактерийг харилцан таних нь трихобластын гадаргуу дээр байрлах тусгай уураг - лектиныг тодорхой нянтай холбох замаар хийгддэг. Бактериас ялгардаг өсөлтийн дааврын нөлөөн дор үндэс үс нь гажигтай байдаг. Эдгээр эрчилсэн эсүүдэд бактери нэвтэрдэг. Эсийн дотор бактери үржиж, бохьны бүрхүүлд бэхлэгдсэн халдварын утас үүсгэдэг. Энэ утас нь үндэс рүү гүн нэвтэрдэг. Бактерийн ийм нэвтрэлтийн үр дүн нь үндэс мезодерм эсийн хүчтэй тархалт бөгөөд энэ нь зангилаа үүсэхэд хүргэдэг. Зангилааны хамгийн гадна хэсэг нь удаан хугацааны туршид меристемийн бүс хэвээр үлдэж, зангилааны дотоод хэсэгт бактери байрладаг. Бактерийн эд нь өөрөө наалдамхай цитоплазмтай, асар олон тооны бактери бүхий жижиг эсүүдээр төлөөлдөг. Бактерийн эд нь үндэсийн төв цилиндртэй холбогдсон судасны багцаар хүрээлэгдсэн байдаг. Тэдгээр. бактери, ургамлын хооронд шим тэжээл солилцох тээврийн систем байдаг. Азот тогтоогч бактери нь ургамлын азотын шим тэжээл, хөрсний үржил шимт сайн нөлөө үзүүлдэг.
Ургамлыг ус, эрдэс бодисоор хангахтай холбоотой үндэсийн физиологийн идэвхийг тодорхой мөөгөнцөртэй симбиотик байдлаар сайжруулж болно. Үндэс, мөөгөнцөрийн ийм ойр дотно харилцан ашигтай симбиоз юм микориза.Микориза нь ихэнх судасны ургамалд мэдэгддэг. Цөөн хэдэн цэцэглэдэг гэр бүлүүд үүнийг үүсгэдэггүй эсвэл ховор тохиолддог, жишээлбэл, байцаа, шанага.
Микориза мөөг нь судасны ургамлын эрдэс тэжээлд гол үүрэг гүйцэтгэдэг. Фосфорыг шингээх, шилжүүлэхэд микориза чухал үүрэг гүйцэтгэдэг.
Хоёр талын тээвэрлэлтийн үед (фосфатыг мөөгөнцөрөөс ургамал руу шилжүүлэх, нүүрс усыг эсрэг чиглэлд шилжүүлэх) ургамал ба мөөгөнцрийн сийвэнгийн мембран хоорондын апопластик бүсэд алдагдал, шингээлт үүсдэг.
Микориза мөөг нь ихэвчлэн ургамалд байдаггүй P-ийн эх үүсвэрийг ашигладаг. Микориза нь ургамалд Zn, Mn, Cu шингээхэд хувь нэмэр оруулдаг. Хөрсөн дэх эдгээр элементүүд нь сул хөдөлгөөнтэй байдаг тул тэдгээрийн шавхагдсан бүсүүд нь үндэс үсний эргэн тойронд хурдан үүсдэг. Микориза мөөгөнцөрийн гифийн сүлжээ нь үндэс бүрээс хэдэн см-ээр үргэлжилдэг бөгөөд ингэснээр үндэс шингээх гадаргууг нэмэгдүүлдэг.
Дараах төрлийн микориза байдаг.
1) ectomycorrhiza: мөөгөнцөр нь үндэсний бүх үзүүрийг зузаан бүрхэвчээр бүрхэж, эс хоорондын зайд нэвтэрдэг;
2) эндомикориза: мөөгөнцөр нь эсийг эзэлдэг.
Ectomycorrhiza нь мод, бут сөөг (нарс, бургас, beech) тодорхой гэр бүлийн шинж чанартай байдаг. Ectomycorrhiza нь залуу, хараахан гүн гүнзгийрээгүй үндэс дээр үүсдэг. Ectomycorrhizal үндэс нь богино, салаалсан, хавдсан харагддаг.
Ийм үндэст язгуур үсний хөгжил дарангуйлж, оройн меристем болон үндэсийн малгайны эзэлхүүнийг багасгаж болно. Үсний үндэсийн үүргийг hyphae гүйцэтгэдэг.
Endomycorrhizal үндэс нь хэлбэрийн хувьд энгийн үндэстэй төстэй боловч тэдгээрээс бараан өнгөөр ялгаатай байдаг. Мөөгөнцрийн hyphae нь үндэс бор гадаргын эсүүдэд нэвтэрч, цэврүүт (хавдар), арбускулууд (мод шиг мөчрүүд) үүсгэдэг. Тиймээс эндомикоризаг ихэвчлэн vesicular-arbuscular гэж нэрлэдэг. Тодорхой нөхцөлд мөөгөнцрийн hyphae нь эсүүдээр "шингээх" боломжтой байдаг. Үндэс дээр үндэс үс нь хадгалагдана. Энэ бол микоризагийн хамгийн түгээмэл төрөл юм. Энэ нь бүх судасны ургамлын ойролцоогоор 80% -д, тухайлбал өвслөг ургамал, түүний дотор үр тариа; бут сөөг, мод (алим мод, агч ...). Жишээлбэл, залуу улиас ургамалд цэврүүт-арбускуляр эндомикориза үүсдэг ба хөгшрөх тусам бүрээстэй эктомикориза үүсдэг.
Ericaceae heather нь эрикоид-арбутоид микоризагаар тодорхойлогддог. Мөөгөнцөр нь үндэс орчимд гифийн хүчтэй хуримтлал үүсгэдэг бөгөөд нимгэн хажуугийн гифүүд нь үндэс бор гадаргын эсүүдэд нэвтэрдэг. Эдгээр микориза мөөгөнцөр нь хөрсний хоруу чанарыг эрс бууруулж, эдгээр ургамлыг хүчиллэг, шим тэжээл багатай хөрсөнд суулгах боломжийг олгодог.
Өөр нэг төрөл байдаг - цахирмаа эндомикориза. Байгаль дахь цахирмаа цэцгийн үр нь зөвхөн тохирох мөөгөнцөр байгаа тохиолдолд соёолдог.
Ерөнхийдөө байгалийн нөхцөлд микоризийн холбоо нь үл хамаарах зүйл биш харин дүрэм юм.
Хамгийн эртний ургамлын үлдэгдэлд хийсэн судалгаагаар эндомикориза нь орчин үеийн ургамлаас багагүй тохиолддог болохыг харуулж байна. Энэ нь хувьслын явцад микориза үүсэх нь ургамлыг газар нутгийг колоничлох боломжийг олгосон хүчин зүйлүүдийн нэг байж магадгүй гэж таамаглах боломжтой болсон. Энэхүү колоничлолын үед хөрсний хөгжил сул байгааг харгалзан үзэхэд микориза мөөг нь ургамлын эрдэс тэжээлийн үндэс суурь болсон гэж үзэж болно. Микориза бүхий орчин үеийн анхдагч ургамлууд нь ядуу хөрсөнд амьд үлдэх магадлал өндөр байдаг.
Микотроф төрлийн тэжээллэг ургамал нь техноген орчинд хүнд металлын агууламжид илүү тэсвэртэй байдаг.
Халуун орны ширэнгэн ойн ургамлын үндэсийн талаар хэдэн үг хэлье. Тулгуур, самбар шиг тулгуур үндэстэй харьцуулахад илүү түгээмэл байдаг. Үндэс нь модны дээд үзүүрийг наасан хавтанг санагдуулам. Ургамал үндэс нь их биен дээр гарч доошоо ургадаг гэнэтийн үндэс байдаг бол банзны үндэс үүсэх нь хөрсний гадаргаас доош шууд ургадаг хажуугийн үндэстэй холбоотой байдаг. Хажуугийн үндэс нь модны их биетэй зэргэлдээх хөндлөн огтлолын гурвалжин хэлбэртэй босоо ургалт үүсгэдэг.
Банзан модны үндэс нь ихэвчлэн ижил модны бусад модноос илүү хатуу байдаг бөгөөд ихэвчлэн нимгэн холтосоор хучигдсан байдаг. Эдгээр үндэснүүдийн хөндлөн огтлолд харагдах өсөлтийн цагиргуудаас харахад эхэндээ тэдгээр нь хэвийн, бөөрөнхий үндэстэй байдаг боловч дараа нь дээд талд нь маш хүчтэй зузааралт үүсдэг. Модон дээрх банзны үндэсийн тоо 1-10 хооронд хэлбэлздэг боловч ихэнх тохиолдолд гурав ба түүнээс дээш байдаг. Халуун оронд банзны үндэс үүсэх нь намгархаг ойд ургадаг мод, чийг муутай хөрсөнд онцгой ажиглагддаг. Ихэнхдээ самбар хэлбэрийн үндэс нь үйлдвэрлэлийн зориулалтаар ашиглагддаг. Тиймээс Индонезид ургадаг Koompassia excelsa модны банз шиг үндсийг хоолны ширээ хийхэд ашигладаг.
ЗУГТАЛ
Найлзуурууд нь үндэс шиг ургамлын гол вегетатив эрхтэн юм. Найлзуурууд нь агаарын тэжээлийн функцийг гүйцэтгэдэг, бусад хэд хэдэн функцийг гүйцэтгэдэг бөгөөд метаморфоз хийх чадвартай.
Ургамлын өөрчлөгдөөгүй найлзуурууд нь тэнхлэгийн хэсэг - ишнээс бүрдэнэ; навч, нахиа. Найлзуурыг үндэснээс нь ялгах гол гадаад шинж чанар нь найлзууруудын навчит байдал юм.
Хажуугийн эрхтнүүд (салбар, навч гэх мэт) үүсэхтэй холбоотой ишний хэсэг - зангилаа.Зэргэлдээх зангилааны хоорондох ишний хэсэг завсрын зангилаа.Навч ба ишний хоорондох дотоод өнцөг навчны суга.Найлзууруудын давтагдах элемент бүр (навч, нахиа бүхий зангилаа, навчны суга ба завсрын зангилаа) үүсдэг. метамер.Тиймээс найлзуурууд нь хэд хэдэн метамеруудаас бүрддэг, i.e. метамерик бүтэцтэй.
20-р зууны 90-ээд оны эхээр Америкийн эрдэмтэд зангилаа ба завсрын тухай ойлголтын оронд - гэсэн ойлголтыг санал болгов. ишний элемент.Ишний элемент нь навчны primordia ирмэгийн хоорондох ишний хэсэг юм. Одоохондоо зангилаа байхгүй байгаа найлзуурын оройд илэрсэн. Тэдгээр. "Үүдэл элемент" гэсэн шинэ ойлголт нь "метамер" гэсэн ойлголттой тохирч байна.
Найлзууруудын оройн меристемийн бүтэц нь үндэстэй харьцуулахад илүү төвөгтэй байдаг. Үнэн хэрэгтээ найлзууруудад оройн меристем нь анхдагч эд эсийн эсийг үүсгэдэг төдийгүй навчны анхдагч ба суганы нахиа үүсэхэд оролцдог бөгөөд дараа нь хажуугийн найлзуурууд үүсдэг. Энэ меристем нь эх малгай шиг бүтцээр хамгаалагдаагүй.
Оройн төрлүүд нь үндэстэй ижил байдаг: нэг төрлийн, бүсчилсэн, шаталсан. Давхаргын тоо нь үндэснээс их байна.
Хувьслын хувьд анхдагч моноинициал орой. Энэ төрлийн оройн жишээг гэзэгнээс олж болно. Ганц том сунасан оройн эс нь хүчтэй вакуолжсон байдаг. Энэ нь түүний гадаргуутай параллель чиглэлд хуваагдана (хамгийн гаднах гадаргуугаас бусад). Хамгийн ойрын гаралтай эсүүд нь маш их вакуолжсан байдаг боловч хуваагдах явцад нягт протопласт бүхий жижиг эсүүд гарч ирдэг. Ийм эсүүд нь ишний оройн захын дагуу байрладаг бөгөөд навчны анхдагч нахиа тавьдаг.
Ихэнх цэцэглэдэг ургамлын найлзууруудын ургамлын орой нь эсүүдийн хуваагдлын хавтгайд ялгаатай хоёр бүс - биет хэлбэртэй байдаг. Туник(лат. - бүрхүүл) нь антиклиналь (гадаргуутай перпендикуляр) хуваагддаг эсийн гаднах давхаргыг багтааж, гадаргуугийн өсөлтийг хангадаг. Хүрээ(лат. - их бие) - дээлний доорх эсийн төв бүлэг, өөр өөр хавтгайд хуваагдаж, найлзууруудын үндсэн эзэлхүүнийг бүрдүүлдэг. Хувцасны бие болон давхарга бүр өөрийн гэсэн анхны үсэгтэй байдаг. Хувцасны эхний үсгийг тэнхлэгийн дагуу төвд байрлуулна. Хоёр талт малтмал нь ихэвчлэн хоёр давхаргатай, монокотууд нь нэг давхаргатай, ихэвчлэн хоёр давхаргатай байдаг.
Туник бүрхүүлийн тухай ойлголт нь оройн меристем дэх эсийн байршил, өсөлтийг ойлгоход хувь нэмэр оруулдаг бол зэргэлдээх хэсгүүдийн өөр өөр эсийн бүсийг таних нь меристемийн эд, эрхтнийг ялгахад оруулсан хувь нэмрийг илрүүлэхэд тусалдаг. зураг авалт. Оройн меристемд янз бүрийн цитологийн шинж чанартай бүсүүд байдаг цитогистологийн бүсчлэл.
Корпус гэж нэрлэгддэг эсийн бүсэд тохирдог төв эх эсийн бүс.Энэ бүсийн эсүүд их вакуолжсан байдаг бөгөөд энэ нь митозын үйл ажиллагааны харьцангуй бага хувьтай холбоотой байдаг. Төвийн эх эсийн бүс нь захын меристемээр хүрээлэгдсэн байдаг бөгөөд энэ нь хэсэгчлэн дээлээс, хэсэгчлэн корпусаас үүсдэг. Протодермийн гистоген нь дээлний гаднах давхаргаас үүсдэг. Захын меристем нь оройн эхний үсгийн хажуугийн деривативууд болон төвийн эх эсүүдээс үүсдэг. Гол меристем нь төв эх эсүүдийн захын дагуух давхаргад хуваагдсаны үр дүнд үүсдэг. шилжилтийн бүс.Захын бүсийн эсүүд нь нягт протопласттай тул митозын өндөр идэвхжилтэй байдаг.
Ялангуяа өндөр идэвхжилтэй зарим газарт навчны булцуу (эсвэл анхдагч булцуу) тавьдаг.
Шинэ навчны примордиа үүсэх нь эсийн хуваагдлын чиглэл өөрчлөгдсөнтэй холбоотой бөгөөд үүний өмнө цитоплазмын захын давхарга дахь микротубулын чиглэл өөрчлөгддөг. Осмосын даралт нь мөн primordia өсөлтөд нөлөөлдөг.
Идэвхтэй өсөлтийн үед найлзууруудын оройн меристем нь навчны анхдагчийг маш хурдан үүсгэдэг тул зангилаа ба завсрын зангилаа нь эхлээд ялгаатай байдаггүй. Аажмаар навчны давхаргын хоорондох хэсгүүд ургаж, сунаж, завсрын хэлбэрийг авч эхэлдэг. Үүний зэрэгцээ навчны хавсралтын хэсгүүд нь зангилаа үүсдэг.
Навчны үндсэн хэсгүүд гарч ирэх доор вакуолизаци ажиглагдаж байна. Энэ нь анхдагч бор гадаргын болон гүний ялгарах эхлэлийг харуулж байна. Анхан шатны бор гадаргын болон гүн дэх хүчтэй вакуолизаци нь үлдэгдэл меристем гэж нэрлэгддэг меристемийн бүс байгааг илтгэнэ. Энэ нь илүү ялгаатай эдүүдийн дунд хадгалагдсан оройн меристемийн үлдэгдэл юм шиг харагдаж байна. Үлдэгдэл меристем дэх навчны үндэс тавигдсан газрын доор уртааш хуваагдлын тусламжтайгаар дараа нь өргөн эсүүд ургахгүйгээр эхний прокамбиумын бага зэрэг сунасан эсүүд үүсдэг.
Хөгжлийн дараагийн үе шатанд үлдэгдэл меристемд илүү олон прокамбын хэлхээ ялгардаг. Ишний өгөгдсөн түвшний бүх судасны багцууд үүссэний дараа үлдэгдэл меристем нь фасцикуляр паренхимд хуваагдана. Зангилаанууд дээр үлдэгдэл меристемийн хэсэг нь навчны ангархай паренхим болдог.
Прокамбиумыг тасралтгүй цагираг эсвэл тусдаа хэлхээнд хийж болно. Хэрэв прокамби нь тасралтгүй цагирагт оршдог бол a цацраггүй (боодолгүй) төрлийн ишний бүтэц.Хэрэв прокамбиумыг тус тусад нь тавьсан бол a ишний бүтцийн цацрагийн төрөл.Тиймээс анхдагч дамжуулагч эдүүд нь ишний хэсэгт янз бүрийн хэлбэрээр байрладаг: эхний тохиолдолд тасралтгүй цагираг, хоёр дахь тохиолдолд үндсэн паренхимийн хэсгүүдээр тусгаарлагдсан тусдаа дамжуулагч багцууд - үндсэн цацрагууд.
Барьцааны судасны багцад анхдагч флоем нь прокамбийн судлын гадна хэсэгт, анхдагч ксилем нь дотоод хэсэгт илэрдэг. Флоемын дараагийн ялгаа нь төв рүү чиглэн явагддаг: флоемын шинэ элементүүд ишний төвд ойртож гарч ирдэг. Xylem нь эсрэг чиглэлд ялгаатай байдаг - төвөөс зугтах замаар.
Бүхэл бүтэн шон буюу ишний төв хэсэг нь дамжуулагч эдүүд, гол (заримдаа энэ нь нурж унадаг), перициклээс бүрдэнэ. Перициклийн гадна талд анхдагч бор гадаргын хэсэг байдаг. Анхдагч бор гадаргын найрлагад (мөн энэ нь захын меристемээс үүсдэг) хлоренхим, заримдаа шүүрлийн элементүүдийг агуулдаг. Захын меристемийн хамгийн гадна давхарга нь үндэстэй адил экзодерм үүсгэдэггүй, харин судал хэлбэрээр эсвэл тасралтгүй цагирагт (ишний бүтцийн төрлөөс хамааран: багцалсан эсвэл багцгүй) байрладаг колленхимд хуваагддаг. Анхдагч бор гадаргын хамгийн дотоод давхарга нь эндодерм юм. Гэхдээ ишний хувьд энэ нь үндэстэй адил зохицуулалтын функцийг гүйцэтгэдэггүй. Цардуулын үр тариа эндодермэд хуримтлагддаг тул үүнийг цардуултай бүрхүүл гэж нэрлэдэг. Заримдаа энэ нь кальцийн оксалат талстыг агуулдаг.
Перицикл нь меристемийн идэвхээ хурдан алдаж, ишний бүтцийн төрлөөс хамааран тасралтгүй давхарга эсвэл судалтай байдаг склеренхим болон хувирдаг.
Цөм нь нимгэн ханатай паренхим эсүүдээр төлөөлдөг бөгөөд тэдгээрт нөөц шим тэжээл хуримтлагддаг. Талст, салиа гэх мэтээр дүүрсэн идиобласт эсүүд байдаг. Цөмийн захын хэсгийг гэж нэрлэдэг перимедулляр бүс(Грекээр peri - эргэн тойронд, medulla - гол). Перимедулляр бүсийн эсүүд нь жижиг хэмжээтэй бөгөөд тэдний амин чухал үйл ажиллагааг удаан хугацаанд хадгалж байдаг.
Монокотуудад анхны төрөл нь ишний бүтцийн багцын төрөл, i.e. прокамбиум нь утаснуудад байрладаг. Гэхдээ хоёрдогч меристем - камби хэзээ ч үүсдэггүй тул хоёрдогч дамжуулагч эдүүд байдаггүй. Өтгөрүүлсэн ч гэсэн иш нь зөвхөн анхдагч эдээр үүсдэг. Монокотуудын зузаан иш (алга, зуун настын) нь үндсэн паренхимийн эсийн хуваагдал, суналтын улмаас үүсдэг. Энэ өсөлтийг сарнисан хоёрдогч өсөлт гэж нэрлэдэг.
Монокотуудын хувьд анхдагч бор гадаргын болон төв цилиндр нь дикотуудын адил тодорхой хил хязгаараар тусгаарлагддаггүй бөгөөд эпидермисийн доор стела дээш өргөгддөг.
Монокотууд нь ишний бүх хэсэгт хаалттай судаснуудтай тархсан байдаг сарнисан хэлбэрийн ишний бүтцээр тодорхойлогддог. Завсрын завсрын суналтын явцад флоем ба ксилемийн анхдагч элементүүд (протофлоем ба протоксилем) сунаж, устдаг бөгөөд үүний үр дүнд багцын ксилемийн тал дээр том агаар агуулсан хөндий үүсдэг. Иймээс лабораторийн судалгаагаар эрдэнэ шишийн ишний хөндлөн огтлолын байнгын бэлдмэлийн багцын төрлүүдийг судлахад боловсорч гүйцсэн судаснуудад хоёр өргөн метаксилем судас, агаарын хөндий, метафлоем багтдаг нь тодорхой байв. Багц бүр нь склеренхимийн эсийн доторлогоотой байдаг.
Монокотуудад ишний гол хэсэг нь үхэж, түүний оронд хөндий үүсдэг. Тодорхой зангилаатай ийм хөндий ишийг нэрлэдэг сүрэл.Соломина нь ишний захад цагираг үүсгэдэг механик эд эсийн хүчтэй хөгжлөөр тодорхойлогддог. Боловсорч гүйцсэн ургамалд хлоренхим устаж, сүрэл нь склеренхимийн онцлог шинж чанартай алтан шаргал өнгөтэй болдог. Ерөнхийдөө монокотууд нь ишний хүчтэй склерациар тодорхойлогддог бөгөөд бүх эд эсийн 30 хүртэлх хувь нь склеренхим байдаг ба амьд паренхимийн эсийн мембраны хэсэгчилсэн lignification байж болно.
Дикотын ишний хоёрдогч өсөлт нь камбиумын үйл ажиллагааны үр дүн юм. Камбиум нь найлзууруудын өсөлтийн эхний жилийн төгсгөлд гарч ирдэг бөгөөд ишний өтгөрөлтөд хувь нэмэр оруулдаг. Камби нь судасны багц дахь прокамбиумаас, хэсэгчлэн фасцикуляр паренхимаас үүсдэг. Үүссэн газраас хамааран камбиумын хэсгүүдийг фасцикуляр ба фасцикуляр хоорондын камби гэж нэрлэдэг. Фасцикуляр хоорондын бүсийн ирээдүйн камбын эсүүд нь cambial үйл ажиллагаа эхлэхээс өмнө тодорхойлогддог. Камбиум нь эхлээд фасцикулийн ойролцоох эсүүдэд үүсдэг ба дараа нь тархины судсанд гүн тархдаг. Үүний үр дүнд фасцикуляр ба фасцикуляр камбиум нь ишний тасралтгүй камбын цагираг үүсгэдэг.
Хоёрдогч өсөлтийн гурван төрөл байдаг.
1) Aristolochia chircason төрөл.
Эхнийх нь ишний бүтцийн цацрагийн төрөл юм. Анхдагч судасны багцууд нь өргөн гол цацрагаар тусгаарлагддаг. Багцлагдсан камби нь хоёрдогч дамжуулагч эдийг үүсгэдэг ба фасцикуляр хоорондын камби нь цацрагийн паренхимийг үүсгэдэг. Энэхүү цацрагийн бүтэц хадгалагдан үлджээ.
2) Helianthus наранцэцгийн төрөл.
Бүтцийн цацрагийн төрөл нь мөн анхных юм. Гэхдээ хоёрдогч дамжуулагч эдүүд нь фасцикуляр ба фасцикуляр хоорондын камбиумаас бүрддэг. Fascicular cambium-аас үүссэн хоёрдогч судасны багцууд нь анхдагчаас жижиг хэмжээтэй, багцын дээгүүр перициклик гаралтай механик эд байхгүй гэдгээрээ ялгаатай байдаг. Камбийн удаан хугацааны нөлөөгөөр хоёрдогч багцууд улам бүр нэмэгдсээр байна. Тэд ургаж, бие биетэйгээ нийлж, хоёрдогч дамжуулагч эдүүдийн тасралтгүй цагираг үүсгэдэг.
3) Тилиа линден төрөл.
Эхний төрөл нь ишний бүтцийн цацраггүй төрөл юм. Нийтлэг cambial цагираг нь хоёрдогч дамжуулагч эдийг үүсгэдэг. Гэхдээ үе үе камби нь паренхимийн эсүүдийг үүсгэдэг бөгөөд энэ нь хоёрдогч туяанд хуваагддаг.
ишний бүтэц олон наст ургамал
Модлог, бутлаг хоёр талт ургамлууд, түүнчлэн шилмүүст ургамлуудад хоёрдогч өтгөрөлт олон жилийн турш үргэлжилж, зарим зүйлийн голч нь хэдэн метр диаметртэй байдаг. Зузаан нь удаан хугацааны туршид ажилладаг камбиум ба хэсэгчлэн феллоген - хоёрдогч хажуугийн меристемийн үйл ажиллагаатай холбоотой байдаг. Байр зүйн хувьд олон наст модлог ургамлын ишний гурван үндсэн хэсгийг ялгаж болно: холтос, мод, яс. Холтос ба модны хоорондох хил нь камбиумын дагуу урсдаг.
Камби эсүүд нь ердийн меристем эсүүдээс ялгаатай нь үргэлж вакуолжсан байдаг. Камбын эсийн цитоплазмын төлөв байдал улирал солигдох тусам өөрчлөгддөг.
Камбын бүс буюу камбийн цагирагт камбын эсийн хоёр хэлбэр олддог: босоо чиглэлд сунасан булны хэлбэртэй эсүүд (эхний үсэг) ба хэвтээ чиглэлд сунасан бөмбөрцөг эсүүд - туяаны эхний үсэг. Хэрэв булны эсүүд нь ердийн хэвтээ эгнээнд байрладаг бол камбиумыг шаталсан камби гэж нэрлэдэг. Хэрэв ээрмэл хэлбэртэй эсүүдийг байрлуулах ийм дараалал байхгүй бол камби нь шатлалгүй байна.
Урт шугамын камби нь урт шугамын мод, урт шугамын хоёрдогч флоемыг үүсгэдэг. Давхаргагүй камби нь шатлалгүй мод ба шатлалгүй хоёрдогч флоемыг үүсгэдэг. Хувьслын хувьд урт шугамын мод, флоем нь давхар биш модыг бодвол илүү нарийн мэргэшсэн гэж үздэг.
Ишний зузааралтаар камбиумын давхаргын тойрог мөн нэмэгддэг. Энэ нь радиаль чиглэлд cambial эсийн хуваагдлаас болж боломжтой юм.
Камбын эсүүдийн дунд эхний давхарга нь тусгаарлагдсан байдаг. Хуваагдсаны үр дүнд түүний эсүүд дотоод, дараа нь дотор нь дериватив үүсгэдэг гадагш чиглэсэн чиглэл. Үүнтэй ижил хугацаанд 2-3 эсийг дотогшоо, зөвхөн нэг эсийг гадна талд нь байрлуулна. Үүний үр дүнд модноос илүү мод үүсдэг.
Цацрагийн эсүүд (эхний үсэг) нь үндсэн паренхимийн туяа үүсгэдэг: үндсэн, голоос бор гадаргын сунадаг; болон хоёрдогч үндсэн цацраг нь богино байдаг, учир нь хожим үүссэн цацрагийн эсүүдээс үүсдэг.
Гол дам нуруу нь нэг эгнээ, олон эгнээ байж болно; нэгэн төрлийн (жишээ нь, ижил эсүүдээс бүрддэг) ба гетероген (цацраг нь бүтцийн хувьд өөр өөр эсүүдийг агуулдаг).
Гол цацрагийн утга : энэ нь унтаа үед (жишээлбэл, өвлийн улиралд) үүрэг гүйцэтгэдэг шим тэжээл (цардуул, липид) хадгалах газар юм.
Гол цацрагууд нь ус ба эрдэс давсыг апопласт дагуух ксилемээс камби ба хоёрдогч флоем руу, шим тэжээлийн симпластын дагуу эсрэг чиглэлд радиаль дамжуулалтыг хангадаг.
Эцэст нь хийн солилцоо нь эс хоорондын зайгаар явагддаг.
Мод
Мод нь камбигаар үүсгэгддэг бөгөөд төвөөс зугтах чиглэлд ургадаг. Учир нь Манай бүсэд камби нь үе үе ажилладаг: хаврын эхэн үед ажиллаж эхэлдэг бөгөөд зуны сүүлээр үйл ажиллагаагаа зогсоодог, дараа нь модонд төвлөрсөн давхарга харагдана. Хавар, өсөн нэмэгдэж буй улирлын эхэн үед эрт мод үүсдэг. Түүний судаснууд нь өргөн, нимгэн ханатай, их хэмжээний ус зөөх чадвартай. Өсөлтийг эхлүүлэх, ялангуяа шинэ эсийн хэмжээг нэмэгдүүлэх (жишээлбэл, навчны эсийг хөгжүүлэх) ус хэрэгтэй. Хожуу мод агуулсан цөөхөн хөлөг онгоц. Тэд илүү нарийхан, зузаан байдаг. Тухайн жилийн модны эхэн үеэс сүүл рүү шилжих шилжилт нь аажмаар явагддаг. Гэсэн хэдий ч, эрт мод нь нэг жилийн хожуу модны хил ирэх жилүргэлж хурц бөгөөд хөндлөн огтлолын хэсэгт тод харагддаг. Үүний үр дүнд жилийн өсөлт (жилийн цагираг эсвэл жилийн давхарга) үүсдэг.
Жилийн (жилийн) өсөлтөд хөлөг онгоцны байршлын дагуу дараахь төрлийн модыг ялгадаг.
Хэрэв савнууд нь ихэвчлэн эрт модонд байрладаг бол хожуу модонд тэдгээр нь байхгүй эсвэл зөвхөн царс, үнсэн мод зэрэг жижиг савнууд байрлуулсан бол энэ нь цагираг хэлбэртэй мод.
Хэрэв хөлөг онгоцнууд жилийн өсөлтийн туршид жигд тархсан боловч хожуу модны голч нь багасдаг бол энэ нь тархай бутархай судасны мод.Хус, агч, улиас зэрэгт олддог.
Эдгээр төрлийн модны хооронд шилжилтийн хэлбэрүүд бас байдаг.
Мод нь дамжуулагч, механик, хадгалалтын функцийг гүйцэтгэдэг тул ксилемийн элементүүдээс гадна склеренхим хэлбэрийн механик эд, үндсэн паренхим хэлбэрийн хадгалалтын эдийг агуулдаг. Бүх анатомийн элементүүдийн бүтэц, тэдгээрийн тоо, байршил нь янз бүрийн ургамлын модыг тодорхой ангилал зүйн бүлгийн онцлог шинж чанартай болгодог.
Жишээлбэл, хувьслын хувьд илүү дэвшилтэт ургамал нь тусгай механик эдтэй (склеренхим). Хувьслын анхдагч хоёр талт (замбага) нь модны утасгүй байдаг. Энэ тохиолдолд механик үүргийг трахеидууд гүйцэтгэдэг.
Зарим ургамлын модонд трахеидээс libriform руу шилжих шилжилтийн хэлбэрүүд байдаг (libriform нь модны тусгай механик элементүүд бөгөөд зузаан хүрэн бүрхүүлтэй төгсгөлд нь үзүүртэй прозенхим эсүүдээс бүрддэг). Мөн клойсон libriform байдаг. Зузаан босоо мембран бүхий анхны булны хэлбэртэй эс нь тусдаа богино амьд эсүүдэд хуваагддаг. Ийм гистологийн элементүүд нь морфологийн болон үйл ажиллагааны хувьд модлог паренхимд ойрхон байдаг.
Модны паренхим нь хадгалах, хэсэгчлэн дамжуулах үүргийг гүйцэтгэдэг. Паренхимийн эсүүд амьд байдаг. Модны паренхимийг хэвтээ ба босоо эгнээнд байрлуулна. Хэвтээ эгнээ нь үндсэн цацрагийг үүсгэдэг. Бид тэдний онцлог шинж чанарыг аль хэдийн авч үзсэн. Босоо эгнээ нь модлог паренхимийг өөрөө үүсгэдэг.
Хэрэв модны паренхим нь жилийн өсөлтийн туршид тархсан байвал энэ сарнисан паренхим.Линден, царс, лийр модны хувьд ердийн зүйл.
Хэрэв модны паренхим нь хөлөг онгоцны эргэн тойронд байрладаг бол энэ паратрахеаль паренхим.Үнс, агч мод.
Жилийн өсөлтийн гаднах хил дээр байрладаг терминал паренхим.Жишээ нь: бургас, шинэс, замбага мод.
Түүний бүтцийн онцлог нь цусны судас байхгүй байх явдал юм. Гуурсан хоолойн элементүүд нь цооролтгүй бөгөөд дамжуулагч болон механик үүрэг гүйцэтгэдэг трахеидоор төлөөлдөг.
Шилмүүст трахеидууд нь торус бүхий том хилтэй нүхээр тодорхойлогддог. Трахейд нь урт, үзүүртэй үзүүртэй. Эрт трахеидууд нь том, нимгэн ханатай байдаг. Хожим нь трахеидууд нь радиаль хавтгайрсан, зузаан ханатай байдаг.
Гимносперм ба дикотуудын мод нь бүтцийн хувьд ихээхэн ялгаатай болохыг бид харж байна. Тиймээс үүнийг хоёр үндсэн бүлэгт хуваадаг. Гимноспермийн модыг зөөлөн, дикот модыг хатуу мод гэж ангилдаг. Эдгээр "хатуу" ба "зөөлөн" гэсэн нэр томъёо нь нягтрал, хатуулгийн хэмжээгээр хязгаарлагдах албагүй. Ялгаанууд нь бид аль хэдийн хэлсэнчлэн бүтцийн онцлогтой холбоотой байдаг.
Налуу эсвэл муруй хонгилд реактив мод үүсдэг. Түүний үүсэх нь салбар, их бие нь налуу байх үед үүсдэг ачааллыг эсэргүүцэх хандлагаас үүдэлтэй, i.e. түүний үүсэх нь ургамлын эдгээр хэсгүүдийг шулуун болгох үйл явцтай холбоотой юм.
Реактив модны хөгжилд нөлөөлдөг хамгийн чухал хүчин зүйлүүд нь таталцлын хүч ба дотоод өсөлтийг өдөөгч бодисын тархалт юм.
Реактив мод нь анатомийн болон химийн хувьд ердийн модноос ялгаатай. Реактив модны эсүүд нь лигнификация, ханын өтгөрөлтийг их хэмжээгээр хийдэг. Шилмүүст модны хувьд энэ нь хүрээлэн буй эдээс илүү нягт, бараан өнгөтэй байдаг. Түүний трахеидууд нь ердийн модныхоос богино байдаг.
Модны цацраг идэвхт чанар нь гадна давхаргаас цөм рүү чиглэн буурдаг. Их биений ёроолоос орой хүртэл модны бохирдол бага зэрэг нэмэгддэг.
Холтос, найлзуурууд дахь цацраг идэвхт бодисын хангалттай өндөр агууламж нь тэдгээрийн гаднах агаарын бохирдлыг илтгэнэ.
Холтос
Энэ нэр томъёонд камбиумаас гадуурх бүх эдүүд орно. Энэ нь аль хэдийн хоёрдогч холтос юм, учир нь. нь хоёрдогч меристемээр үүсдэг - камби (анхдагч меристемийн эсийг ялгах явцад үүсдэг анхдагчаас ялгаатай).
Хоёрдогч бор гадаргын найрлагад хоёрдогч флоем, склеренхим (баст утас ба чулуурхаг эс), үндсэн паренхим орно. Механик элементүүдийн багц гэж нэрлэдэг хатуу баст,дамжуулагч элементүүд ба үндсэн паренхим - зөөлөн баст.
Нууцлаг эсүүд болон давирхайн суваг нь хоёрдогч бор гадаргын хэсэгт байрладаг. Хоёрдогч бор гадаргын гол үүрэг нь хуванцар бодисыг дамжуулах, түүнчлэн хамгаалалтын функц юм.
Хоёрдогч флоем нь их биений эзэлхүүний хоёрдогч ксилемээс хамаагүй бага хувийг эзэлдэг. Үүнийг нэгдүгээрт, камби нь ксилемээс бага флоем үүсгэдэг, хоёрдугаарт, хуучин, ажиллахгүй флоемыг аажмаар буталдагтай холбон тайлбарладаг.
Тэнхлэгийн эрхтний тойрог нэмэгдсэний улмаас паренхимийн туяа өргөжиж, гурвалжинтай төстэй бөгөөд орой нь камби руу чиглэсэн байдаг. Ийм паренхимийг өргөссөн (лат. dilatatio - тэлэлт) гэж нэрлэдэг. Паренхимийн ийм зохион байгуулалт нь ишний зузаан ургах тусам баст урагдахаас сэргийлдэг.
Шилмүүст ургамалд хоёрдогч флоем нь илүү энгийн бүтэцтэй байдаг. Гимноспермийн баст гистологийн элементүүд жигд тархсан бөгөөд мэдээжийн хэрэг давирхайн суваг байдаг.
Феллогений гаднах бүх эдүүд (хамгийн дотоод үйсэн камбий гэсэн үг) үйсэн эсүүд субэринжих үед ус, эрдэс бодис хүлээн авахаа болино. Кортексийн үлдэгдэл (паренхим ба флоемын үхсэн эсүүд) бүхий эдгээр бүх перидермүүд нь гаднах (гадна) бор гадаргыг бүрдүүлдэг. Камби ба феллогений хамгийн дотоод давхаргын хоорондох бор гадаргын амьд хэсгийг дотоод кортекс гэж нэрлэдэг.
Тиймээс бид "Эд" сэдвээр олж авсан мэдлэг дээрээ үндэслэн ишний тэнхлэгийн эрхтэн болох ишний анатомийн бүтцийг судалж үзсэн.
Одоо найлзууруудын хажуугийн эрхтэн болох навчны бүтцийг авч үзье.
ХУУДАС
Навч нь дээд ургамлын фотосинтезийн гол эрхтэн юм. Навчны бүтэц, түүний үүрэг нь хоорондоо нягт холбоотой байдаг.
Фотосинтезийн урвалын нийт тэгшитгэлээс:
CO 2 + H 2 O chl-l → hv (CH 2 O) n + O 2
1) навч CO 2, усны эх үүсвэр хэрэгтэй гэж бид дүгнэж болно; 2) навч нь шингээхэд тохирсон байх ёстой нарны эрчим хүчтэд хлорофиллтэй байх ёстой; 3) O 2 нь урвалын бүтээгдэхүүний нэг хэлбэрээр ялгарах болно; 4) нүүрс усыг нөөцөд хадгалах эсвэл ургамлын бусад хэсэгт тээвэрлэх ёстой. Жагсаалт бол энэ бүх шаардлагыг хангасан маш нарийн мэргэжлийн байгууллага юм.
Ургамлын эрхтэн болох навчны бүх хувьсал нь гэрлийг хамгийн сайн ашиглахад дасан зохицох хөгжлийн замыг дагасан.
Микрофилийн хувьслын шугамын дагуу навчийг ринофит төрлийн эртний ургамлын теломын ургалт гэж үздэг бөгөөд тэнхлэгийн эрхтэний бүтцийн бүтцийг хадгалдаг.
Макрофил хувьслын шугамын дагуу навч нь хавтгайрч, теломуудыг нэг хавтгайд байрлуулж, дараа нь нэг бүхэлд нь нэгтгэсний үр дүнд үүссэн. Үүний зэрэгцээ урт хугацааны оройн өсөлт, салбарлах чадвар алдагдсан. Урт хугацааны хувьслын үр дүнд үүссэн навчны морфологи, анатомийн шинж чанарууд юу вэ?
Насанд хүрэгчдийн навч нь ихэвчлэн навчны ир, ишний ишнээс бүрдэнэ. Дэлбээ- навчны ир ба найлзууруудын зангилааны хоорондох навчны нарийхан иштэй хэсэг, түүний тусламжтайгаар навчийг орон зайд чиглүүлж, гэрэлтэй харьцуулахад хамгийн таатай байдлаар байрлуулна. Дэлбээнээр дамжуулан навчны ир ба иш нь холбогддог. Дэлбээнд дамжуулагч ба механик эдүүд сайн хөгжсөн байдаг. Навч унах үед ишний хэсэгт тусгаарлах давхарга үүсдэг.
Навчны хамгийн доод хэсэг, ишний хамт зангилаа гэж нэрлэдэг хуудасны суурь.Навчны суурь нь янз бүрийн хэлбэртэй байж болно. Ихэнхдээ энэ нь жижиг зузааралт шиг харагддаг бөгөөд үүнийг нэрлэдэг навчны дэвсгэр.Ихэнхдээ хуудасны ёроолд байдаг янз бүрийн хэлбэрүүдба хосолсон хажуугийн ургалтын хэмжээ - stipules. Stipules нь навч хэлбэртэй байж болох ба нэмэлт шингээх гадаргуу болж үйлчилдэг. Стипулууд нь зүү хэлбэртэй байж болох ба ургамлын хамгаалалтын үүрэг гүйцэтгэдэг.
Хайрст үлд, хальслаг стипулууд байдаг. Хувьслын хувьд стипулын бууралт ажиглагдаж, хувьслын залуу ургамлуудад тэд эрт унадаг эсвэл огт байхгүй байдаг.
Зарим үр тариа, селөдерей (шүхэр) нь навчны суурь нь ургаж, хаалттай эсвэл нээлттэй хоолой үүсгэдэг - навчны бүрээс. Навчны бүрээс нь завсрын меристемийг удаан хугацаанд хадгалахад хувь нэмэр оруулж, найлзуурыг нэмэлт дэмжлэг болдог.
Навчны гол хэсэг нь навчны ир юм. Навчны дээд ба доод талыг жигд бус гэрэлтүүлсний үр дүнд хавтан нь dorsoventral бүтэцтэй, i.e. навчны дээд талын бүтэц, ховдол, доод тал, нурууны бүтцээс ялгаатай. Навчны дээд талыг ховдол гэж нэрлэдэг, учир нь. бөөрөнд энэ тал нь дотоод бөгөөд найлзуурын тэнхлэг рүү эргэдэг. Нахиа дахь навчны доод, нуруу, тал нь ургамлаас гадагш чиглэсэн байдаг. Тиймээс dorsoventral тэгш хэмтэй навч нь хоёр талт эсвэл хоёр талт эсвэл хоёр нүүртэй байдаг. дикотуудын онцлог.
Радиаль тэгш хэмтэй навч нь тэгш өнцөгт юм. Ийм өтгөрүүлсэн, цилиндр хэлбэртэй навчнууд нь Crassulaceae-д байдаг. Нэг талт, эсвэл нэг талт, нэг талт навч нь үр тариа, сонгины шинж чанартай байдаг.
Санаж үз навчны ирний анатомийн бүтэц.
Үндэс, ишний нэгэн адил навч нь салст бүрхэвч, дамжуулагч, үндсэн эдээс бүрдэнэ. Учир нь Навч нь ихэвчлэн хоёрдогч ургалтгүй байдаг (дэлбээ ба том судлын бага зэрэг ургалтаас бусад), эпидермисийг салст бүрхэвч болгон хадгалдаг.
Стоматууд ихэвчлэн навчны доод талд байдаг. Хоёр талт навчнуудад стоматууд навчны бүх гадаргуу дээр харагдахгүй дарааллаар тархсан байдаг. Монокот болон шилмүүст навчны хувьд тэдгээр нь навчны уртааш тэнхлэгтэй зэрэгцээ эгнээнд тархсан байна. Зарим үр тарианы эпидермисийн хэсэгт мотор эсвэл моторт эсүүд байдаг. Эдгээр нь ердийн эпидермисийн эсүүдээс том хэмжээтэй байдаг. Вакуоль нь бараг бүхэл эсийг эзэлдэг. Тургор алдагдсанаар эдгээр эсүүд агшиж, навчийг нугалж, нугалахад хувь нэмэр оруулдаг.
Навчны ирний үндсэн эдүүдийн ихэнх нь мезофилд байдаг. Мезофилл нь ялгаатай байдаг. Палисад мезофилл нь хавтангийн дээд талд, хөвөн - ёроолд байрладаг. Ийм бүтэцтэй навчнууд нь хоёр нүүртэй эсвэл нурууны хэлбэртэй байдаг.
Хэрвээ палисад мезофилл нь навчны хоёр талд байрладаг бол навч нь нэг нүүртэй эсвэл изоталт байдаг.
Үр тарианы мезофилд палисад ба хөвөн хлоренхим гэж ялгагддаггүй. Мезофилл эсүүд нь багцын эргэн тойронд радиаль байдлаар байрладаг.
Дамжуулагч навчны багцыг судал гэж нэрлэдэг ба судлын салаалсан системийг нэрлэдэг Venation.Навчны ир дэх судасны багцын байршлын онцлог нь дихотом, параллель, нуман, далдуу, зүү хэлбэртэй байж болох янз бүрийн хэлбэрийн венийн судсыг тодорхойлдог.
Дээд салаалсан эрэмбийн судаснууд хоорондоо хэрхэн холбогдож байгаагаас хамааран нээлттэй (дихотом) ба хаалттай судлуудыг ялгадаг. Дихотомийн судалтай үед судлууд нь дихотомоор салаалж, хурц өнцгөөр салж, навчны ирний ирмэг дээр хүрч, хөршийн судалтай хаана ч нийлдэггүй. Хаалттай судалтай бол жижиг судлууд нь анастомозын үр дүнд (гатралт) бүхэл хавтан эсвэл түүний ихэнх хэсгийг нэвчих сүлжээ үүсгэдэг.
Навчнууд нь далдуу, зууван судалтай бөгөөд эдгээр нь ихэвчлэн хоёр талт, дунд, хамгийн том судалтай байдаг. Судас нь анхдагч ксилем ба анхдагч phloem-ээс тогтдог бөгөөд прокамбиумаас үүссэн ба хавсарсан багцад нэгтгэгддэг. Камбиум ихэвчлэн ксилем ба флоемын хооронд үүсдэг боловч энэ нь ажилладаггүй. Тиймээс ксилем ба флоем нь анхдагч юм.
Багцууд нь ишний дамжуулагч системтэй холбогдсон навчинд тасралтгүй систем үүсгэдэг. Иймээс ксилем нь морфологийн хувьд навчны дээд тал руу үргэлж чиглэсэн байдаг бол флоем нь морфологийн хувьд үргэлж доод тал руу чиглэгддэг.
Дунд судал нь хажуугийн жижиг судлуудаар холбогддог. Тэд тус бүр нь бүр жижиг хэмжээтэй холбоотой бөгөөд тэдгээр нь эргээд бүр жижиг хэсгүүдэд хуваагддаг гэх мэт. Дикотууд дахь салаалсан захиалгын тоо 2-оос 5 ба түүнээс дээш хооронд хэлбэлздэг. Хамгийн жижиг мөчрүүд нь хаалттай эсүүдийг үүсгэдэг жижиг талбайнуудмезофилл - арола.
Нээлттэй дихотомийн венийн системд хаалттай эсүүд байдаггүй.
Монокот навчинд (зэрэгцээ венацтай) уртааш судлууд нь энгийн хөндлөн гүүр хэлбэрээр байрладаг жижиг судлууд - комиссын багцуудаар хоорондоо холбогддог. Жижиг судлууд дахь дамжуулагч эдүүдийн тоо аажмаар буурдаг. Багцын төгсгөлд ксилемийн элементүүд нь флоемоос илүү хол зайд үргэлжилдэг. Венийн төгсгөлд байрлах ксилем нь ихэвчлэн гуурсан хоолойн богино элементүүд, флоем нь богино нарийн шигшүүр элементүүд ба том хиймэл эсүүдээс тогтдог.
Мезофилд байрлах жижиг судасны багцууд нь багцын бүрээсийг бүрдүүлдэг нягт атираат эсийн нэг буюу хэд хэдэн давхаргаар хүрээлэгдсэн байдаг. Багцын бүрээс нь паренхим эсвэл склеренхим байж болно. Зарим зүйлийн хувьд фасцикулын бүрээс дэх эсийн ханын суберинизаци ажиглагдаж байгаа нь бүрхүүлийн эсүүд эндодермийн үүрэг гүйцэтгэдэг болохыг харуулж байна.
Колленхим ба склеренхим нь нэг буюу хоёр талдаа том судлууд дээр байрладаг. Том судлуудтай холбоотой эд нь навчны гадаргуугаас дээш гарч, цухуйсан хэсгүүдийг үүсгэдэг. Судасны цухуйсан хэсгүүдийн хооронд байрлах эд эсийн хэсгийг "хос хоорондын бүс" гэж нэрлэдэг (лат. Цутгах нэр нь "коста" - хавирга).
Зузаан навчтай зүйлүүдэд мезофилд янз бүрийн склерейд байдаг бөгөөд тэдгээр нь зайны үүрэг гүйцэтгэдэг.
Үр тарианы навчнууд нь склеренхимийн хүчтэй хөгжлөөр тодорхойлогддог.
Гимноспермийн навчнууд нь ангиоспермийн навчнаас арай бага бүтэцтэй байдаг. Өвлийн улиралд бага температурт дасан зохицох чадвар, хэдэн жил амьдрах чадвар нь анатомийн бүтцэд хэрхэн тусгагдсан болохыг нарс зүүгээр жишээ болгон авч үзье. Зүү нь зузаан ханатай эпидермитэй, хүчтэй зүслэг, гүн шингэсэн стоматай. Зарим шилмүүст ургамлуудын хажуугийн эсүүд (Cupressaceae) нь стоматын хамгаалалтын эсүүд дээр унжсан зүслэг нуруутай байдаг. Стоматууд нь зүүний бүх талд босоо эгнээнд байрладаг. Эпидермисийн доор зузаан ханатай, гиподермисийн хэсэгчлэн бордсон эсийн мембранууд байдаг. Мезофилл нь нугалж байна. Энэ нь давирхайн хэсгүүдийг агуулдаг. Дамжуулагч багцууд нь зүүний төвд байрладаг. Ихэвчлэн хоёр, ховор нэг. Эдгээр багцууд нь склеренхимийн хусуураар холбогддог. Багцууд нь трахеид ба паренхимийн эсүүдээс бүрдсэн цус сэлбэх эдээр хүрээлэгдсэн байдаг. Цус сэлбэх эд (лат. transfusio - цус сэлбэх) нь дамжуулагч багц ба мезофиллийн хооронд ус, шим тэжээлийг тээвэрлэхтэй холбоотой.
Цус сэлбэх эд нь зузаан ханатай эндодермээр хүрээлэгдсэн байдаг. Эндодерм нь ихэвчлэн хөгжлийн эхний үе шатанд Каспар судалтай, хожуу үе шатанд суберик хавтантай байдаг. Боловсорч гүйцсэн төлөвт эндодерм эсүүд хоёрдогч lignified мембрантай байдаг.
Навчны ирний анатомийн бүтэц нь мэдэгдэхүйц уян хатан чанарыг харуулдаг. Ургамлын оршин тогтнох нөхцөл, ургамал дээр навч байрлуулах зэргээс шалтгаалан бүтэц өөрчлөгддөг.
Орчин үеийн ботаникийн хөгжлийн нэг чиглэл болох ургамлын экологийн анатомийг бид аль хэдийн тэмдэглэсэн бөгөөд энэ нь эсийн амьд агуулга - протопласт нь хүрээлэн буй орчны нөхцөл байдлын өөрчлөлтөд хамгийн түрүүнд хариу үйлдэл үзүүлдэг гэж хэлсэн. Тиймээс хойд ургамлын эсүүд бага вакуолжсан, том хлоропласттай байдаг. Стромд олон цардуулын үр тариа байдаг. Хлоропластын тилакоидын систем нь сэрүүн өргөргийн ургамлаас бага хөгжсөн байдаг. Хойд ургамлын митохондрийн тоо 2-2.5 дахин их байдаг. EPR болон Голжийн аппарат илүү хөгжсөн. Хойд ургамлын мезофиллийн эсийн субмикроскопийн бүтцийн ийм шинж чанарууд нь цаг уурын хатуу ширүүн нөхцөлд ургамлын амьд үлдэхэд хувь нэмэр оруулдаг чухал дасан зохицох шинж чанарууд юм.
Чийгийн дутагдалд дасан зохицсон ургамалд хүйтэн, зузаан эсийн мембран, ялангуяа эпидермисийн давхаргад тэмдэглэгдсэн байдаг. Том эсийн эпидермисийн хүчтэй өтгөрүүлсэн гаднах мембран нь эсийн хөндийн хагасыг эзэлдэг. Эпидермисийн дээгүүр зузаан зүсмэл давхарга байдаг. Гадаргуугаас харахад зүслэг нь зузаан лаваар хучигдсан байдаг бөгөөд энэ нь навчны хөхрөлтийг тодорхойлдог.
Стоматууд нь эпидермисийн үсээр гадагшилдаг хотгорт дүрдэг.
Шүүслэг ургамал нь ус хадгалах эдтэй байдаг.
Зөөлөн квиноа (Африк) навчны чийгийг барих сонирхолтой дасан зохицох арга. Дээд ба доод талаас навч нь олон эгнээ байрлуулсан цэврүүт үстэй тасралтгүй давхаргаар хучигдсан байдаг. Энэ давхаргын зузаан нь мезофиллийн зузаанаас ихээхэн давж, амьд эдийг хатахаас хамгаалдаг. Үс нь шөнийн цагаар агаарын чийгийг барьж, хатуу ширүүн байдлыг сайжруулдаг усны горимургамал.
Манай тархай бутархай ойн бүсийн квиноагийн навч, иш нь олон тооны бөмбөлөг хэлбэртэй үсээр хучигдсан байдаг нь нунтаг бүрээстэй мэт сэтгэгдэл төрүүлдэг боловч тэдгээр нь зөөлөн квиноа шиг тийм өтгөн олон эгнээ давхарга үүсгэдэггүй.
Хоёр төрлийн quinoa-ийн навчийг харьцуулах жишээнээс харахад ижил төстэй ангилал зүйн нэгжүүдийн системчилсэн шинж чанарууд нь экологийн өөр өөр нөхцөлд суурьшсан үед тэгш бус хөгжлийг олж авдаг бөгөөд дасан зохицох шинж чанартай байдаг.
Борооны ойн ургамлын навчнууд (жишээ нь, мангас) нь цоолсон навчны ир, урт иштэй байдаг. Илүүдэл ус үргэлж амархан урсдаг. Стоматууд нээлттэй байна. Их хэмжээний агаарын хөндий бүхий палисад даавуу.
Температур, чийгшилээс гадна өөр нэг чухал абиотик хүчин зүйл бол гэрэл бөгөөд ургамал нь бүтцийн өөрчлөлтөд хариу үйлдэл үзүүлдэг.
Нарны шууд тусгалд ургаж буй навчнууд нь сүүдрийн навчнаас жижиг боловч палисад мезофилл хүчтэй хөгжсөний үр дүнд сүүдэр навчнаас зузаан байдаг.
Тиймээс хувьслын явцад тогтсон хүрээлэн буй орчны нөхцөл байдалд дасан зохицох нь тоон зөрүүтэй холбоотой боловч зохион байгуулалтын зарчмуудыг өөрчилдөггүй.
Навчны онтогенез
Навчнууд нь оройн меристемийн хажуугийн ургалт хэлбэрээр жижиг булцуу хэлбэртэй байдаг. Навчны булцуу үүсэхэд зөвхөн дээл төдийгүй биеийн эсүүд оролцдог. Навчны примордиа нь анхны цухуйсан хэсгээс дээшээ конус эсвэл зүү хэлбэртэй ургалт хэлбэрээр ургадаг. Удалгүй навчны үндсэн хэсгүүд оройн меристемийг хүрээлж, түүнийг механик болон амьсгалын явцад ялгаруулдаг дулаанаар хамгаалдаг.
Эрт навч ургах нь ихэвчлэн оройн ба ахиу (ахиу) гэж хуваагддаг. Эхнийх нь primordia-ийн суналт, хоёр дахь нь хажуугийн өсөлттэй холбоотой бөгөөд энэ нь хавтангийн хоёр хэсэг үүсдэг. Үүний дагуу өсөн нэмэгдэж буй приморди нь оройн орой дээр оройн меристем, тэнхлэгийн хажуу тал дээр хоёр захын меристемтэй байдаг. Примордиагийн оройн өсөлт удаан биш юм. Оройн ургалт ба хавтангийн ирмэгүүд нь завсрын өсөлттэй байдаг.
Нийлмэл навчинд захын меристемийн үйл ажиллагаа нь тусдаа төвүүдэд байрладаг бөгөөд тус бүр нь тусдаа ухуулах хуудас үүсгэдэг.
Навчны ирний хөгжил нь хожим нь завсрын өсөлтөөс үүсдэг ишний өсөлтөөс хурдан байдаг.
Хэрэв завсрын суурь ба захын бүсийн хоорондох хил дээр нарийсал үүссэн бол ишний навч үүснэ. Дэлбээний ихэнх хэсэг нь завсрын бүсээс үүсдэг. Хэрэв ийм нарийсалт (интеркаляр болон захын бүсийн суурийн хоорондох хил дээр) үүсэхгүй бол суумал навч үүснэ.
Монокотилет ургамлын навчны үндсэн хэсэгт ахиу меристем бараг үүсдэггүй бөгөөд урт хугацааны туршид завсрын өсөлт нь онцлог шинж чанартай байдаг. Тиймээс навчны ирний гол хэлбэр нь шугаман хэлбэртэй байдаг.
Хавтангийн янз бүрийн давхарга дахь эсийн хуваагдал ба суналтын хурдны ялгаа нь олон тооны эс хоорондын зай, ердийн мезофиллийн бүтэц үүсэхэд хүргэдэг.
Бусад эдүүдийн хөгжил нь дамжуулагч эдүүдийн хөгжилтэй холбоотой байдаг. Прокамби нь навч болон өсөлтийн конусын зэргэлдээх хэсэгт нэгэн зэрэг салж, тасралтгүй судал үүсгэдэг. Энэ нь анхдагч судасны багцыг үүсгэдэг бөгөөд энэ нь үргэлжилсэн бөгөөд навч, ишний хувьд нийтлэг байдаг.
Навчны онтогенез нь навч ба ишний нийтлэг шинж чанарыг нэг бүхэл хэсэг болох найлзуурыг гэрчилдэг.
Нахиа үүссэн цагаас эхлэн навчис үүсэх нахиалах нэмэлт үе эхэлдэг. Үүний зэрэгцээ навчны гадаргуу хэдэн зуун, бүр хэдэн мянган удаа нэмэгддэг. Гадаргуугийн өсөлт нь навчны ихэнх эсийг хувааж, урт, өргөнөөр нь сунгах замаар хийгддэг.
Навчны насжилт нь удамшлын болон цаг уурын хүчин зүйлээс хамаардаг. Сэрүүн уур амьсгалтай навчит мод, бут сөөгний хувьд навчис нахиалах хугацаа 4-5 сар байна. Шилмүүст ургамлын навч нь хоёроос зургаа ба түүнээс дээш жил амьдардаг. Гэхдээ ямар ч тохиолдолд навчны нас нь ургамлын тэнхлэгийн эрхтнүүдийн насжилтаас хамаагүй бага байдаг.
Welwitschia mirabilis нь баруун өмнөд Африкийн чулуурхаг цөлд ургадаг. Энэхүү одой мод нь урт үндэстэй, зузаан, богино их биетэй (50 см өндөр, 1 м хүртэл зузаантай), амьдралын туршид (2000 жил хүртэл) хадгалагддаг хоёр том навчтай. Навчны урт нь 2-3 метр хүрч, ёроолдоо байнга ургаж, дээд хэсэгт нь үхдэг. Velvichia-ийн чийгийн бараг цорын ганц эх үүсвэр бол өтгөн манан бөгөөд чийгийг навчны хоёр тал дахь олон тооны stomata-аар шингээдэг (1 см 2 тутамд 22,200 stomata).
Амьд эд эсийг гэмтээхгүйгээр навчийг мөчрөөс нь идэвхтэй салгах үйлдлийг abscision гэж нэрлэдэг. Модны улирлын навчис уналт нь өдрийн уртын өөрчлөлтөд үзүүлэх хариу урвалын үр дүн юм. Намрын улиралд навч унах нь намар, өвлийн улиралд физиологийн чийгийн дутагдалтай нөхцөлд зайлшгүй шаардлагатай ургамлын ууршилтын гадаргууг ихээхэн бууруулдаг. Навч унасны ачаар цасны жингээс мөчрийг хугалах эрсдэл буурдаг.
Ихэнх навчинд тусгаарлах давхарга үүсэх нь онтогенезийн үед тохиолддог. Энэ бүсэд механик эд эсийн хэмжээ багасдаг. Гуурсан хоолойн элементүүдэд урсахаас өмнө thyls үүсдэг ба калоз нь шигшүүрийн элементүүдэд хуримтлагддаг. Хагарлын үед эсийн мембраны ферментийн устгал үүсдэг бөгөөд энэ нь эсийг бие биенээсээ салгахад хүргэдэг. Мембран дахь өөрчлөлтүүд нь дунд давхаргын цементлэх чадвар алдагдах (хэсэгчлэн түүнээс кальцийг зайлуулсантай холбоотой), целлюлозын мембран өөрөө гидролиз, гуурсан хоолойн цэвэршсэн элементүүдийн тасархай зэрэг орно.
Суберин эсвэл шархны бохь зэрэг хамгаалалтын бодисууд мембран, эс хоорондын зайд хуримтлагдсаны үр дүнд хамгаалалтын давхарга буюу сорви үүсдэг. At модлог ургамалхамгаалалтын давхарга нь хамгаалалтын давхарга дор тавигдаж, ишний бусад хэсгийн перидермтэй тасралтгүй үргэлжилдэг перидермээр солигдоно.
Навч унах нь эсийн мембраныг татан буулгах үйл явцтай холбоотой байх албагүй. Урьдчилсан химийн өөрчлөлтгүйгээр механик салгах нь гацуураас зүү унах үед ажиглагдаж болно.
Навч унасны дараа навчны сорви нь тэдний баглаа сорвитой үлддэг. Цацрагийн сорви- эдгээр нь навч унахаас өмнө навчны ул мөрнөөс навчны иш рүү явсан судасны багцын эвдэрсэн төгсгөлүүд юм.
навчны ул мөр- эдгээр нь ишний дамжуулагч системээс навч хүртэлх мөчрүүд юм. Навчны ул мөр нь ишний боодолтой нийлсэн газраас навч руу гарах хүртэл сунадаг. Нэг навч нь нэг буюу хэд хэдэн навчны ул мөр байж болно.
Учир нь иш ба навч нь филогенетикийн хувьд нийтлэг гарал үүсэлтэй тул навчны ул мөр ба ишний багц хооронд үндсэн ялгаа байхгүй. Холбогдох нэр томъёо нь дүрслэх, байр зүйн утгатай байна.
Навчны ул мөр ишний төв дамжуулагч цилиндрээс навчны суурь руу шилждэг зангилааны бүсэд цилиндрт паренхимийн бүс үүсдэг - навчны завсар эсвэл навчны завсар. Навчны ул мөр, навчны лакунагийн тоо нь ургамал бүрт өөр өөр байдаг.
Ангиоспермийн зангилааны үндсэн төрөл нь гурав ба түүнээс дээш тооны лакунатай нэг юм. Ангиоспермийн хувьслын гол чиглэл нь лакунагийн тоо буурах замаар тодорхойлогддог.
Ургамлын тэнхлэгийн хэсгийн дотоод цөм (иш, үндэс) эсвэл дамжуулагч системийн захад байрлах лакуна, цөм, фасцикуляр хоорондын бүс, дамжуулагч систем ба перицикл. стела.
Стелагийн ангилал нь хөгжлийн анхдагч үе шатанд тэнхлэгийн эрхтнүүдийн дамжуулагч ба дамжуулагч бус эдүүдийн харилцан зохицуулалт дээр суурилдаг. Филогенетикийн хувьд хамгийн анхдагч гэж тооцогддог хамгийн энгийн хэлбэрийн стелад дамжуулагч эд нь ксилемийг тойрсон флоем бүхий тасралтгүй багана үүсгэдэг. Энэ бол protostele юм. Ишний ийм зохион байгуулалт нь биологийн хувьд төгс бус, учир нь дамжуулагч болон суурь эдүүдийн хоорондох холбоо барих гадаргуу бага байна.
Стелийн цаашдын хувьсал нь дамжуулагч эдүүдийн контактын гадаргууг үндсэн хэсгүүдтэй нэмэгдүүлэх замаар явав. Үүнд хоёр аргаар хүрсэн. Нэг тохиолдолд стела нь бор гадаргын зүг гүн ургалт үүсгэдэг бөгөөд хөндлөн огтлолын хувьд од хэлбэртэй байдаг. Энэ бол актинотел юм.
Хоёрдахь, биологийн хувьд илүү ирээдүйтэй хөгжлийн арга бол стелийн төвд паренхимийн цөм үүсэх явдал юм. Энэ бол сифон юм.
Дамжуулагч ба үндсэн эдүүдийн хоорондын нягт холбоо нь навчис хөгжихөд үүсдэг. Илүү их навч, стеле нь үндсэн паренхимаар ханасан байдаг. Стелла нь сүлжээний хэлбэрийг авдаг (Грек сүлжээ - диктуон). Энэ бол диктиостеле юм.
Орчин үеийн цэцэглэдэг ургамлын стеле нь бүр илүү паренхиматжсан байдаг. Дамжуулагч судал ба фасцикуляр хоорондын бүсийн систем болох стела нь eustela (Грекээр eu - сайн) юм.
Монокотуудад навчны ул мөр ишний төвд хүрч, дараа нь зах руу хазайдаг. Тиймээс ишний хөндлөн огтлол дээр боодол нь ямар ч дараалалгүйгээр тараагдсан мэт харагдана. Грек хэлнээс гаралтай Атактос. замбараагүй. Тиймээс монокотуудын стелийн нэр - атактостеле. Тэр онцлогтой өндөр зэрэгтэй parenchymatization, үед навч бүр ул мөр "
Үндэс нь тэнхлэгийн эрхтэн бөгөөд радиаль тэгш хэмтэй бөгөөд оройн меристем хадгалагдаж байвал урт нь ургадаг. Үндэс нь ишнээсээ морфологийн хувьд навчис хэзээ ч харагдахгүй, оройн меристем нь үндсээр бүрхэгдсэн байдгаараа ялгаатай. Үндэс үр удмын ургамалд салаалсан нахиа үүсэх нь перицикл (анхдагч хажуу меристем)-ийн үйл ажиллагааны үр дүнд эндоген (дотоод) хэлбэрээр явагддаг.
Root функцууд:
1) Үндэс нь хөрсөнд ууссан эрдэс бодис бүхий усыг шингээдэг;
2) ургамлыг хөрсөнд бэхлэх зангууны үүргийг гүйцэтгэдэг;
3) шим тэжээлийн сав болж үйлчилдэг;
4) тодорхой органик бодисын анхдагч нийлэгжилтэнд оролцдог;
5) үндэс үр удамд ургамлын нөхөн үржихүйн үүргийг гүйцэтгэдэг.
Үндэс ангилал. Гарал үүслээр нь үндэс нь үндсэн, нэмэлт, хажуугийн гэж хуваагддаг. Үрийн үр хөврөлийн үндэсээс үүссэн үндсийг гэнэ гол; ургамлын бусад эрхтнүүд (иш, навч, цэцэг) дээр үүссэн үндэс гэж нэрлэдэг adnexal. Насанд хүрэгчдийн ургамалд (монокот ба олон хоёр талт ургамал) үндэс систем нь ихэвчлэн (эсвэл зөвхөн) гэнэтийн үндэсээс бүрддэг тул өвслөг ангиоспермийн амьдралд гэнэтийн үндэсийн үүрэг асар их байдаг. Найлзууруудын суурь хэсэгт гэнэтийн үндэс байгаа нь ургамлыг салангид найлзуурууд эсвэл гэнэтийн үндэстэй найлзууруудын бүлэг болгон хуваах замаар зохиомлоор үржүүлэхэд хялбар болгодог.
Залуу ишний хувьд нэмэлт үндэс нь эндоген байдлаар үүсдэг - фасцикуляр хоорондын паренхимийн эсүүдээс, хуучин нь - гол цацрагийн камбиумын ойролцоох паренхимаас, өөрөөр хэлбэл паренхимийн гаралтай камбиас үүсдэг. Монокот ишний шинэ үндэс нь багцыг тойрсон паренхимд үүсдэг.
Хажуу талүндэс нь үндсэн болон нэмэлт үндэс дээр үүсдэг. Тэдний цаашдын салбарлалтын үр дүнд дээд зэрэглэлийн хажуугийн үндэс гарч ирдэг. Ихэнх тохиолдолд салбарлах нь дөрөв, тав дахь дараалал хүртэл тохиолддог.
Нэг ургамлын бүх үндэсийн нийлбэрийг нэрлэдэг үндэс систем.
гарал үүслээр нь:
үндсэн үндэс системүр хөврөлийн үндэсээс үүсдэг бөгөөд хоёр дахь болон дараагийн эрэмбийн хажуугийн үндэс бүхий үндсэн үндэсээр (эхний дараалал) төлөөлдөг. Зөвхөн үндсэн үндэс систем нь олон мод, бут сөөг, нэг наст болон зарим олон наст өвслөг дикотуудад хөгждөг;
гэнэтийн үндэс системиш, навч, заримдаа цэцэг дээр хөгждөг. Үндэсний энэхүү гарал үүслийг илүү анхдагч гэж үздэг, учир нь энэ нь зөвхөн гэнэтийн үндэстэй системтэй дээд спорын шинж чанартай байдаг. Ангиоспермийн нэмэлт үндэсийн систем нь үрнээс үүсдэг бөгөөд үүнд протокорм (үр хөврөлийн булцуу) үүсдэг ба дараа нь түүн дээр гэнэтийн үндэс үүсдэг;
холимог үндэс системДикот болон монокотуудын дунд өргөн тархсан. Үрнээс ургасан ургамалд үндсэн үндэс нь эхлээд үүсдэг боловч түүний өсөлт удаан үргэлжилдэггүй - энэ нь ихэвчлэн эхний ургалтын улирлын намар зогсдог. Энэ үед үндсэн найлзууруудын гипокотил, эпикотил, дараагийн метамерууд, дараа нь хажуугийн найлзууруудын суурь хэсэгт гэнэтийн үндэсийн систем тогтмол хөгждөг. Ургамлын төрлөөс хамааран тэдгээр нь метамеруудын тодорхой хэсгүүдэд (зангилаа, доод ба түүнээс дээш, зангилаа) эсвэл бүхэл бүтэн уртын дагуу эхэлж, хөгждөг.
Үндэс системийн ангилал хэлбэрээр.
Үндсэн үндэс системийг гэж нэрлэдэг чухалүндсэн үндэс нь урт, зузаан нь хажуугийнхаас мэдэгдэхүйц давсан бол;
Үндсэн ба хажуугийн үндэс нь ижил төстэй хэмжээтэй, үндэс системтэй утаслаг. Холимог үндэс систем нь бас байж болно чухал, хэрэв үндсэн үндэс нь бусдаас хамаагүй том бол, утаслагхэрэв бүх үндэс харьцангуй тэнцүү хэмжээтэй бол. Үүнтэй ижил нэр томъёо нь гэнэтийн үндэсийн системд хамаарна.
Нэг үндэс системд үндэс нь ихэвчлэн өөр өөр үүргийг гүйцэтгэдэг. Араг ясны үндэс (дэмжих, хүчтэй, механик эдүүд хөгжсөн), өсөлтийн үндэс (хурдан ургадаг, гэхдээ бага зэрэг салаалсан), хөхөх (нимгэн, богино настай, эрчимтэй салаалсан) байдаг.



































- Хэмжээ: 2 MB
- Слайдын тоо: 36
Илтгэлийн тайлбар ҮНДЭС, ҮНДЭСНИЙ ТОГТОЛЦОО 1. Функци ба хувьслын слайдууд
ҮНДЭС, ҮНДЭСНИЙ СИСТЕМ 1. Үндэсний үүрэг, хувьслын гарал үүсэл. 2. Үндэсний анхдагч бүтэц. 3. Хоёрдогч язгуур өөрчлөлтүүд. 4. Хажуугийн болон нэмэлт үндэс үүсэх. үндэс системүүд. 5. Үндэсийг мэргэшүүлэх, өөрчлөх.
Үндэс нь радиаль тэгш хэмтэй, оройн меристемийн үйл ажиллагааны улмаас урт нь хязгааргүй ургадаг тэнхлэгийн эрхтэн юм. Навч нь үндэс дээр хэзээ ч харагдахгүй бөгөөд оройн меристем нь үргэлж малгайгаар бүрхэгдсэн байдаг. Үндэсний гол үүрэг бол ус, эрдэс бодисыг шингээх, өөрөөр хэлбэл ургамлыг хөрсний тэжээлээр хангах явдал юм. Дээр дурдсан үндсэн функцээс гадна үндэс нь бусад үүргийг гүйцэтгэдэг: тэд хөрсөн дэх ургамлыг бэхжүүлж, босоо ургах, гөлөглөх боломжтой болгодог; үндэс нь янз бүрийн бодисын хоёрдогч синтез (амин хүчил, алкалоид, фитогормон гэх мэт) байдаг; сэлбэг бодисыг үндэст байрлуулж болно; үндэс нь бусад ургамлын үндэс, хөрсний бичил биетэн, мөөгөнцөртэй харилцан үйлчилдэг.
Үндэс нь хөрсний гадаргуу дээр тархсан ринофитуудын теломуудаас үүссэн. Хувьслын явцад эдгээр теломын зарим мөчрүүд хөрсөнд гүнзгийрч, үндсийг үүсгэсэн.
Үндэс нь илүү төгс хөрсний тэжээлд тохирсон байдаг. Үндэс үүсэх нь тэдний бүх бүтцийн гүн гүнзгий бүтцийн өөрчлөлттэй хамт байв. Тэд тусгай эд эстэй. Хөрсний бодисыг шингээх үүргийг залуу үндэсийн төгсгөлүүд гүйцэтгэж эхлэв. Тэд гадаргуу дээр амьд эсийг хадгалдаг. Эдгээр эсүүд нь үйл ажиллагааны хувьд хамгийн чухал үндэс эд болох үндэслэг ишийг бүрдүүлдэг.
Хөрсний бодисыг шингээх үүргийг залуу үндэсийн төгсгөлүүд гүйцэтгэж эхлэв. Тэд гадаргуу дээр амьд эсийг хадгалдаг. Эдгээр эсүүд нь үйл ажиллагааны хувьд хамгийн чухал үндэс эд болох үндэслэг ишийг бүрдүүлдэг. Цаашилбал, хувьслын явцад шингээх үндэс гадаргуу нь гурван хүчин зүйлийн нөлөөгөөр нэмэгдсэн: 1) элбэг дэлбэг салаалж, олон тооны шингээгч төгсгөл үүсэх; 2) үндэсийн байнгын өсөлт, сорох төгсгөлүүдийн хөрсний шинэ хэсгүүдэд шилжих; 3) үндэс үс үүсэх.
Үндэс ургалт нь өтгөн хөрсөнд байдаг тул түүний оройн меристем хамгаалагдсан байх ёстой. Оройн меристемийг эх малгайгаар гэмтээхээс хамгаалсан. Үндэс үүсэх нь уур амьсгалын хуурайшилт ихэссэнтэй холбоотой байв. Илүү хуурай уур амьсгалтай болж эхэлснээр хуурай газрын ургамал субстрат дээр наалдаж, ус, шим тэжээлийг шингээхэд хүргэсэн. Гэсэн хэдий ч хувьслын явцад янз бүрийн ургамлын үндэсийн бүтэц нь ишнийхээс бага өөрчлөгдсөн. Энэ нь хөрсний орчны нөхцөл агаараас илүү тогтвортой байдагтай холбоотой юм. Тиймээс үндэс нь зугтахаас хамаагүй хожуу гарч ирсэн ч илүү "консерватив" эрхтэн гэж тооцогддог. Үндэс үүсэх нь чухал ургамлын ароморфоз юм. Түүний ачаар ургамал хуурай хөрсийг эзэмшиж, том найлзуурыг бий болгож чадсан.
Үндэс малгайны амилопластын хүндийн хүчний хариу үйлдэл. Хөдлөх статолитууд тоглодог чухал үүрэгбосоо үндэс өсөлтийг хангадаг фитогормоны градиентийг бий болгоход.
Улаан буудайн суулгацын үндэсийн бүтэц (Triticum aestivum): А - үндэсийн бүтцийн диаграмм; B - rhizodermal болон exoderm эсийн ялгаа. 1 - дамжуулалтын бүс, 2 - сорох бүс, 3 - сунах бүс, 4 - хуваах бүс, 5 - үндэс үс, 6 - эх малгай.
Үндэсний хөндлөн огтлол (a - нэг яст, б - хоёр талт ургамал)
Анхдагч бор гадаргын гажигнаас үүсдэг. Түүний гол масс нь нимгэн мембран бүхий амьд паренхим эсүүдээс тогтдог. Тэдгээрийн хооронд үндэс тэнхлэгийн дагуу сунгасан эс хоорондын зайны систем үүсдэг. Хий (CO 2) нь эс хоорондын зайгаар эргэлддэг. Хий нь бор гадаргын болон rhizodermis-ийн эсүүдэд эрчимтэй бодисын солилцоог хангахад шаардлагатай байдаг. Бор гадаргын эсүүд дэх энергийн солилцоо нь хэд хэдэн чухал үүргийг гүйцэтгэхэд зайлшгүй шаардлагатай: 1) бор гадаргын эсүүд нь үндэслэг хальсыг хуванцар бодисоор хангаж, өөрөө бодисыг шингээх, дамжуулахад оролцдог; 2) бор гадаргын янз бүрийн бодисууд нийлэгжиж, дараа нь бусад эдэд шилждэг; 3) нөөц бодисууд нь бор гадаргын эсүүдэд хуримтлагддаг; 4) бор гадаргын дотор микориза үүсгэдэг мөөгөнцрийн гиф ихэвчлэн байдаг.
Эндодерм эсүүд хөгжлийн гурван үе шатыг дамждаг. Шингээх бүсэд эндодерм эхний шатанд байна. Түүний эсийн радиаль хананы дунд Каспарийн туузууд үүсдэг. Каспарийн туузууд нь эсийн мембранаар, өөрөөр хэлбэл апопласт дагуух бодисын хөдөлгөөнийг хаадаг. Хоёр дахь үе шатыг хажуугийн үндэсийн бүсэд ажиглаж болно. Үүний зэрэгцээ эсийн мембраны дотор талд нимгэн суберин хавтан гарч ирдэг. Гэсэн хэдий ч эндодерм нь нимгэн ханатай бие даасан эсүүдийг агуулдаг тул уусмалыг чөлөөтэй дамжуулдаг. Эндодермийн хөгжлийн гурав дахь үе шат нь монокотуудын үндэс хэсэгт ажиглагдаж болно. Түүний эсийн дотоод болон радиаль хана нь их хэмжээгээр өтгөрдөг. Хөндлөн хэсгүүдэд эдгээр эсүүд тах хэлбэртэй байдаг. Шалгах цэг байхгүй. Зузаан ханатай эндодерм нь дамжуулагч эдийг хамгаалж, үндэсийн бат бөх чанарыг нэмэгдүүлдэг.
Каспарийн бүс нь ус нэвтэрдэггүй хаалт бөгөөд усыг апопластаас гадагшлуулж, эндодермийн эсийн мембранаар дамжин симпласт руу урсдаг.
Хөрсний үндэс рүү ус урсах: ус нь эндодермисэд хүрэх хүртэл апопласт ба симпластын дагуу хөдөлж болно. Апопласт дагуу цаашдын хөдөлгөөн хийх боломжгүй юм.
Үндэсний төв цилиндрийн бүтцийн янз бүрийн хэлбэрүүд (анхдагч бүтэц): А-диархи, В-триархи, С-тетрархи, Г-полиархи. А-В төрөлхоёр талт ястны шинж чанар, G - олон монокотод байдаг. 1 - анхдагч бор гадаргын талбай, 2 - анхдагч флоем, 3 - анхдагч ксилем.
Үндэс анхдагчаас хоёрдогч бүтэц рүү шилжих 4 үе шатыг ялгаж салгаж болно: 1) анхдагч флоем ба ксилемийн хэсгүүдийн хоорондох камбиумын харагдах байдал; 2) перициклээр феллоген үүсэх; 3) анхдагч царцдасыг урсгах; 4) дамжуулагч эдүүдийн радиаль байрлалыг барьцаагаар өөрчлөх.
Анхдагчаас хоёрдогч үндэс бүтэц рүү шилжих, 11 - хоёрдогч холтос, 12 - феллоген, 13 - феллем.
Вандуйны үндэс дэх дамжуулагч эдүүдийн анхдагч ялгах схем 1 - ксилемийн төв рүү чиглэсэн ялгаа, 2 - эпидермис, 3 - анхдагч бор гадаргын, 4 - эндодерми, 5 - ксилемийн анхны ялгагдах элементүүд, 6 - ялгагдаагүй ксилемийн элементүүд, 7 - анхны ялгаатай флоем элементүүд , 9 - оройн меристем, 10 үндэстэй малгай.
Монокотуудын үндэс дэх хоёрдогч өөрчлөлт. Морлог ургамлын дийлэнх нь амьдралынхаа эцэс хүртэл анхдагч үндэс бүтцийг хадгалдаг. Гэсэн хэдий ч үндэсийн олон элементүүд нь лигнификацияд ордог. Модон монокотуудад (алга, дракаена, юкка) меристемийн давхарга нь паренхимийн эсүүд эсвэл перициклээс гаралтай эх бор гадаргаас үүсдэг. Үүнээс хаалттай дамжуулагч багцын эгнээ үүсдэг. Энэхүү дамжуулагч багцын дараа анхдагч бор гадаргын паренхимын захын хэсэгт боловсролын эдийн шинэ давхарга гарч ирдэг. Энэхүү меристемийн давхарга нь шинэ цуврал судасны багцыг үүсгэдэг. Тиймээс үндэс нь өтгөрдөг.
Адал үндэс нь ургамлын янз бүрийн эрхтэн дээр - иш, навч, үндэс дээр үүсдэг. Үүдэн дээр үүссэн шинэ үндэсийг ишний үндэс, үндэс дээр үүссэнийг үндэс гэж нэрлэдэг. Хажуугийн болон нэмэлт үндэс нь эндоген гаралтай, өөрөөр хэлбэл дотоод эдэд тавигддаг.
Хажуугийн үндэс эхлэл нь перицикл эсийн хуваагдалаас эхэлдэг. Энэ тохиолдолд стелийн гадаргуу дээр меристемийн булцуу үүсдэг. Меристемийн сүрьеэгийн хэд хэдэн эсийн хуваагдлын дараа хажуугийн үндэс гарч ирдэг. Энэ нь өөрийн оройн меристем, бүрээстэй. Хажуугийн үндэсийн нахиа ургаж, эхийн үндэсийн анхдагч холтосыг нэвтэлж, гадагшаа хөдөлдөг. Ихэвчлэн хажуугийн үндэс нь ксилемийн элементүүдийн эсрэг үүсдэг. Тиймээс тэдгээрийг үндэс дагуу тогтмол уртааш эгнээнд байрлуулна. Тэдгээр нь шингээлтийн бүсэд эсвэл арай өндөр байдаг. Хажуугийн үндсийг тавих нь акропет хэлбэрээр, өөрөөр хэлбэл үндэсийн ёроолоос орой хүртэл явагддаг.
Адвентицийн үндэс нь ихэвчлэн меристемийн үйл ажиллагаа явуулах чадвартай эдэд байрладаг: перицикл, камби, феллоген. Хажуугийн (болон гэнэтийн) үндэс нь эндоген үүсэх нь дасан зохицох чадвартай байдаг. Хэрэв мөчир нь оройд явагдсан бол хөрсөнд үндэс нь урагшлахад хэцүү байх болно.
Хөвдний үндэс систем дэх хоёр талт салаалалт (Lycopodium clavatum) 1 - хамгийн нимгэн үндэсийн изотомын дихотом салбарлалт
Микориза: А - эктотроф царс микориза, В, С - эндотрофын цахирмаа цэцгийн микориза. Люпин үндэс дээрх зангилаанууд