Կողային արմատների ձևավորում. Արմատը, նրա գործառույթները. Արմատների և արմատային համակարգերի տեսակները
Արմատը բույսերի ստորգետնյա առանցքային տարրն է, որը նրանց ամենակարեւոր մասն է, հիմնական վեգետատիվ օրգանը։ Արմատի շնորհիվ բույսը ֆիքսվում է հողում և պահվում այնտեղ իր կյանքի ողջ ընթացքում, ինչպես նաև ապահովվում է դրանում պարունակվող ջրով, հանքանյութերով և սննդանյութերով։ Կան արմատների տարբեր տեսակներ և տեսակներ: Նրանցից յուրաքանչյուրն ունի իր առանձնահատկությունները. Այս հոդվածում մենք կքննարկենք արմատների գոյություն ունեցող տեսակները, արմատային համակարգերի տեսակները: Կծանոթանանք նաև նրանց բնորոշ հատկանիշներին։
Որո՞նք են արմատների տեսակները:
Ստանդարտ արմատը բնութագրվում է թելանման կամ նեղ գլանաձև ձևով: Բազմաթիվ բույսերում, բացի հիմնական (հիմնական) արմատից, զարգացած են նաև արմատների այլ տեսակներ՝ կողային և պատահական։ Եկեք ավելի սերտ նայենք, թե ինչ են դրանք:
հիմնական արմատը
Այս բույսի օրգանը զարգանում է սերմի բողբոջային արմատից։ Միշտ կա մեկ հիմնական արմատ (բույսերի արմատների մյուս տեսակները սովորաբար հոգնակի են): Այն մնում է գործարանում ողջ կյանքի ցիկլի ընթացքում։
Արմատը բնութագրվում է դրական գեոտրոպիզմով, այսինքն՝ ձգողականության շնորհիվ այն խորանում է ենթաշերտի մեջ՝ ուղղահայաց դեպի ներքև։
պատահական արմատներ
Adventitious կոչվում են բույսերի արմատների տեսակները, որոնք ձևավորվում են իրենց մյուս օրգանների վրա: Այդ օրգանները կարող են լինել ցողուններ, տերևներ, ընձյուղներ և այլն։ Օրինակ՝ հացահատիկային կուլտուրաներն ունեն այսպես կոչված առաջնային պատահական արմատներ, որոնք դրված են սերմի ծիլերի ցողունում։ Զարգանում են սերմի բողբոջման գործընթացում հիմնական արմատի հետ գրեթե միաժամանակ։
Գոյություն ունեն նաև արմատների տերևային (առաջացած տերևների արմատակալման արդյունքում առաջացած), ցողունային կամ հանգուցային (առաջացած կոճղարմատներից, վերգետնյա կամ ստորգետնյա ցողունային հանգույցներից) և այլն, ստորին հանգույցներում առաջանում են հզոր արմատներ, որոնք կոչվում են. օդային (կամ օժանդակ):
Վատ արմատների տեսքը որոշում է բույսի վեգետատիվ վերարտադրության ունակությունը:
Կողային արմատներ
Կողային կոչվում են արմատներ, որոնք առաջանում են որպես կողային ճյուղ։ Նրանք կարող են ձևավորվել ինչպես հիմնական, այնպես էլ պատահական արմատների վրա: Բացի այդ, դրանք կարող են ճյուղավորվել կողային արմատներից, ինչի արդյունքում ձևավորվում են ավելի բարձր կարգի կողային արմատներ (առաջին, երկրորդ և երրորդ):
Խոշոր կողային օրգանները բնութագրվում են լայնակի գեոտրոպիզմով, այսինքն, դրանց աճը տեղի է ունենում գրեթե հորիզոնական դիրքում կամ հողի մակերեսի անկյան տակ:
Ի՞նչ է արմատային համակարգը:
Արմատային համակարգ կոչվում է արմատների բոլոր տեսակներն ու տեսակները, որոնք ունի մեկ բույս (այսինքն՝ դրանց ամբողջականությունը): Կախված հիմնական, կողային և պատահական արմատների աճի հարաբերակցությունից՝ որոշվում է դրա տեսակը և բնավորությունը։
Արմատային համակարգերի տեսակները
Եթե հիմնական արմատը շատ լավ զարգացած է և նկատելի է մեկ այլ տեսակի արմատների մեջ, դա նշանակում է, որ բույսն ունի ձողային համակարգ։ Հանդիպում է հիմնականում երկշաքիլ բույսերում։
Այս տեսակի արմատային համակարգը բնութագրվում է հողի մեջ խորը բողբոջումով: Այսպիսով, օրինակ, որոշ խոտերի արմատները կարող են թափանցել 10-12 մետր խորություն (շուշ, առվույտ): Ծառերի արմատների ներթափանցման խորությունը որոշ դեպքերում կարող է հասնել 20 մ-ի։
Եթե պատահական արմատներն ավելի ցայտուն են, զարգանում են մեծ քանակությամբ, իսկ հիմնականը բնութագրվում է դանդաղ աճով, ապա ձևավորվում է արմատային համակարգ, որը կոչվում է թելքավոր։

Որպես կանոն, որոշ խոտաբույսեր նույնպես բնութագրվում են նման համակարգով. Չնայած այն հանգամանքին, որ մանրաթելային համակարգի արմատները չեն թափանցում այնքան խորը, որքան ձողային համակարգի արմատները, նրանք ավելի լավ են հյուսում իրենց հարակից հողի մասնիկները: Բազմաթիվ չամրացված թփերի և կոճղարմատավոր խոտեր, որոնք կազմում են առատ մանրաթելային բարակ արմատներ, լայնորեն օգտագործվում են ձորերը, լանջերին հողերը և այլն ամրացնելու համար: Լավագույն տորֆային խոտերը ներառում են առանց խարույկը, խարույկը և այլն:

փոփոխված արմատներ
Բացի վերը նկարագրված բնորոշներից, կան արմատների և արմատային համակարգերի այլ տեսակներ: Դրանք կոչվում են փոփոխված:
պահեստային արմատներ
Պաշարները ներառում են արմատային մշակաբույսեր և արմատային պալարներ:
Արմատային մշակաբույսը հիմնական արմատի խտացումն է՝ դրա մեջ սննդանյութերի նստվածքի պատճառով: Նաև ցողունի ստորին հատվածը մասնակցում է արմատային բերքի ձևավորմանը։ Բաղկացած է հիմնականում պահեստային հիմքի հյուսվածքից: Արմատային մշակաբույսերի օրինակներ են մաղադանոսը, բողկը, գազարը, ճակնդեղը և այլն:

Եթե խտացած պահեստային արմատները կողային և պատահական արմատներ են, ապա դրանք կոչվում են արմատային պալարներ (կոններ): Մշակված են կարտոֆիլում, քաղցր կարտոֆիլում, դալիաներում և այլն։
օդային արմատներ
Սրանք օդային մասում աճող կողային արմատներ են։ Ներկայացրե՛ք թվով արեւադարձային բույսեր. Ջուրն ու թթվածինը ներծծվում են օդից։ Հասանելի է արևադարձային բույսերի մեջ, որոնք աճում են անբավարարության պայմաններում հանքանյութեր.

շնչառական արմատները
Սա մի տեսակ կողային արմատներ է, որոնք աճում են դեպի վեր՝ բարձրանալով ենթաշերտի, ջրի մակերևույթից: Արմատների նման տեսակներ առաջանում են չափազանց խոնավ հողերի վրա, ճահճային պայմաններում աճող բույսերում։ Նման արմատների օգնությամբ բուսականությունը ստանում է օդից պակասող թթվածինը։
Աջակցող (տախտակաձև) արմատներ
Այս տեսակի ծառերի արմատները բնորոշ են խոշոր տեսակներին (հաճարենի, կնձնի, բարդի, արևադարձային և այլն), դրանք եռանկյունաձև ուղղահայաց աճեր են, որոնք առաջանում են կողային արմատներից և անցնում հողի մակերեսի մոտ կամ վերևում։ Դրանք նաև կոչվում են տախտակաձև, քանի որ նման են ծառին հենված տախտակների։
Ծծող արմատներ (haustoria)
Սա մագլցող բույսերի ցողունի վրա զարգացող լրացուցիչ պատահական արմատների տեսակ է: Նրանց օգնությամբ բույսերը որոշակի հենակետին ամրանալու և վեր բարձրանալու (հյուսելու) հատկություն ունեն։ Նման արմատները հասանելի են, օրինակ, համառ ֆիկուսում, բաղեղում և այլն:

Շարժվող (կծկվող) արմատներ
Բնութագիր բույսերին, որոնց արմատը հիմքում կտրուկ կրճատվել է երկայնական ուղղությամբ։ Օրինակ կարող են լինել բույսերը, որոնք ունեն լամպ: Շարժվող արմատները լամպերին և արմատային մշակաբույսերին ապահովում են հողի մեջ որոշակի անցք: Բացի այդ, դրանց ներկայությունը որոշում է վարդերների (օրինակ, խտուտիկի մեջ) ամուր տեղավորումը գետնին, ինչպես նաև ուղղահայաց կոճղարմատի և արմատային օձի ստորգետնյա դիրքը:
Միկորիզա (սնկի արմատ)
Միկորիզան բարձրագույն բույսերի արմատների սիմբիոզն է (փոխշահավետ համակեցություն) սնկային հիֆերով, որոնք հյուսում են դրանք՝ որպես արմատային մազեր: Սնկերը բույսերին ապահովում են ջրով և դրա մեջ լուծված սննդանյութերով։ Բույսերն իրենց հերթին սնկերին ապահովում են նրանց կենսագործունեության համար անհրաժեշտ օրգանական նյութերով։
Mycorrhiza-ն բնորոշ է շատ բարձր բույսերի, հատկապես փայտային բույսերի արմատներին:
բակտերիալ հանգույցներ
Սրանք փոփոխված կողային արմատներ են, որոնք հարմարեցված են ազոտը ամրագրող բակտերիաների հետ սիմբիոտիկ համակեցության համար: Հանգույցների առաջացումը տեղի է ունենում երիտասարդ արմատների ներթափանցման շնորհիվ: Նման փոխշահավետ համատեղ կյանքը բույսերին թույլ է տալիս ստանալ ազոտ, որը բակտերիաները օդից փոխանցում են իրենց համար մատչելի ձևի: Մյուս կողմից, բակտերիաներին տրվում է հատուկ միջավայր, որտեղ նրանք կարող են գործել՝ չմրցելով բակտերիաների այլ տեսակների հետ: Բացի այդ, նրանք օգտագործում են նյութեր, որոնք առկա են բուսականության արմատներում:

Բակտերիալ հանգույցները բնորոշ են հատիկաընդեղենի ընտանիքի բույսերին, որոնք լայնորեն օգտագործվում են որպես մելիորանտներ ցանքաշրջանառության ժամանակ՝ հողերը ազոտով հարստացնելու նպատակով։ Ազոտ ֆիքսող լավագույն բույսերը համարվում են արմատային հատիկները, ինչպիսիք են կապույտ և դեղին առվույտը, կարմիր և արսափինը, եղջյուրավոր մորեխը և այլն։
Բացի վերը նշված կերպարանափոխություններից, կան նաև այլ տեսակի արմատներ, ինչպիսիք են հենակետային արմատները (օգնում են ամրացնել ցողունը), ցողունային արմատները (օգնում են բույսերին չընկղմվել հեղուկ ցեխի մեջ) և արմատների ծծողները (ունեն պատահական բողբոջներ և ապահովում են վեգետատիվ բազմացում):
Ֆիլոգենետիկորեն արմատն առաջացել է ավելի ուշ, քան ցողունը և տերևը՝ կապված բույսերի ցամաքում կյանքի անցնելու հետ և հավանաբար առաջացել է արմատանման ստորգետնյա ճյուղերից։ Արմատը չունի ոչ տերևներ, ոչ էլ բողբոջներ՝ դասավորված որոշակի հերթականությամբ։ Բնութագրվում է երկարության գագաթային աճով, կողային ճյուղերը առաջանում են ներքին հյուսվածքներից, աճի կետը ծածկված է արմատային գլխարկով։ Արմատային համակարգը ձևավորվում է բույսի օրգանիզմի ողջ կյանքի ընթացքում։ Երբեմն արմատը կարող է ծառայել որպես սննդանյութերի մատակարարման վայրի տեղ: Այս դեպքում այն փոփոխվում է։
Արմատների տեսակները
Հիմնական արմատը գոյանում է բողբոջային արմատից՝ սերմերի բողբոջման ժամանակ։ Ունի կողային արմատներ։
Ցողունների և տերևների վրա զարգանում են պատահական արմատներ։
Կողային արմատները ցանկացած արմատների ճյուղեր են:
Յուրաքանչյուր արմատ (հիմնական, կողային, պատահական) ունի ճյուղավորվելու հատկություն, ինչը զգալիորեն մեծացնում է արմատային համակարգի մակերեսը, և դա նպաստում է հողի մեջ բույսի ավելի լավ ամրապնդմանը և նրա սնուցման բարելավմանը:
Արմատային համակարգերի տեսակները
Գոյություն ունեն արմատային համակարգերի երկու հիմնական տեսակ՝ արմատային արմատը, որն ունի լավ զարգացած հիմնական արմատ, և թելքավոր։ Թելքավոր արմատային համակարգը բաղկացած է մեծ թվով պատահական արմատներից՝ նույն չափերով։ Արմատների ամբողջ զանգվածը բաղկացած է կողային կամ պատահական արմատներից և նման է բլթի։
Բարձր ճյուղավորված արմատային համակարգը ստեղծում է հսկայական կլանող մակերես: Օրինակ,
- ձմեռային տարեկանի արմատների ընդհանուր երկարությունը հասնում է 600 կմ-ի.
- արմատային մազերի երկարությունը՝ 10000 կմ;
- Արմատների ընդհանուր մակերեսը 200 մ 2 է։
Սա շատ անգամ ավելի մեծ է, քան վերգետնյա զանգվածի մակերեսը։

Եթե բույսն ունի հստակ արտահայտված հիմնական արմատ, և զարգանում են պատահական արմատներ, ապա ձևավորվում է խառը տիպի արմատային համակարգ (կաղամբ, լոլիկ):
Արմատի արտաքին կառուցվածքը. Արմատի ներքին կառուցվածքը
Արմատային գոտիներ

արմատային գլխարկ
Արմատն իր ծայրով երկարում է, որտեղ տեղակայված են կրթական հյուսվածքի երիտասարդ բջիջները։ Աճող մասը ծածկված է արմատի գլխարկով, որը պաշտպանում է արմատի ծայրը վնասից և հեշտացնում է արմատի տեղաշարժը հողում աճի ընթացքում։ Վերջին ֆունկցիան իրականացվում է արմատի գլխարկի արտաքին պատերի լորձով ծածկվելու հատկության շնորհիվ, ինչը նվազեցնում է արմատի և հողի մասնիկների միջև շփումը։ Նրանք նույնիսկ կարող են հրել հողի մասնիկները: Արմատի գլխարկի բջիջները կենդանի են, հաճախ պարունակում են օսլայի հատիկներ։ Կափարիչի բջիջները մշտապես թարմացվում են բաժանման շնորհիվ: Մասնակցում է դրական գեոտրոպիկ ռեակցիաներին (արմատների աճի ուղղությունը դեպի Երկրի կենտրոն)։

Բաժանման գոտու բջիջները ակտիվորեն բաժանվում են, այս գոտու երկարությունը նույնը չէ տարբեր տեսակների և նույն բույսի տարբեր արմատներում։
Բաժանման գոտու հետևում կա երկարացման գոտի (աճի գոտի): Այս գոտու երկարությունը չի գերազանցում մի քանի միլիմետրը։
Երբ գծային աճն ավարտվում է, սկսվում է արմատների ձևավորման երրորդ փուլը՝ դրա տարբերակումը, ձևավորվում է բջիջների տարբերակման և մասնագիտացման գոտի (կամ արմատային մազիկների և կլանման գոտի): Այս գոտում արդեն առանձնանում են էպիբլեմայի արտաքին շերտը (ռիզոդերմա)՝ արմատային մազիկներով, առաջնային կեղևի շերտը և կենտրոնական գլան։
Արմատային մազերի կառուցվածքը
Արմատային մազիկները արմատը ծածկող արտաքին բջիջների խիստ երկարաձգված ելքեր են: Արմատային մազերի քանակը շատ մեծ է (200-ից մինչև 300 մազ 1 մմ2-ում): Նրանց երկարությունը հասնում է 10 մմ-ի։ Մազերը ձևավորվում են շատ արագ (խնձորի ծառի երիտասարդ տնկիներում 30-40 ժամվա ընթացքում): Արմատային մազերը կարճատև են: Նրանք սատկում են 10-20 օրվա ընթացքում, իսկ արմատի երիտասարդ հատվածում աճում են նորերը։ Սա ապահովում է հողային նոր հորիզոնների զարգացումը արմատներով։ Արմատը շարունակաբար աճում է՝ ձևավորելով արմատային մազիկների ավելի ու ավելի նոր տարածքներ։ Մազերը կարող են ոչ միայն կլանել նյութերի պատրաստի լուծույթները, այլև նպաստել հողի որոշ նյութերի տարրալուծմանը, այնուհետև կլանել դրանք։ Արմատի այն հատվածը, որտեղ սատկել են արմատային մազերը, որոշ ժամանակ կարողանում է ջուր կլանել, բայց հետո ծածկվում է խցանով և կորցնում այդ ունակությունը։
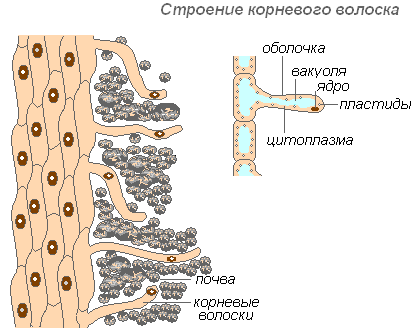
Մազերի պատյանը շատ բարակ է, ինչը հեշտացնում է սննդանյութերի կլանումը։ Գրեթե ամբողջ մազի բջիջը զբաղեցնում է վակուոլը, որը շրջապատված է ցիտոպլազմայի բարակ շերտով։ Միջուկը գտնվում է բջջի վերևում: Բջջի շուրջ ձևավորվում է լորձաթաղանթ, որը նպաստում է արմատային մազերի սոսնձմանը հողի մասնիկներով, ինչը բարելավում է նրանց շփումը և բարձրացնում համակարգի հիդրոֆիլությունը: Կլանումը հեշտացնում է արմատային մազերով թթուների (ածխածնային, խնձորաթթու, կիտրոն) արտազատումը, որոնք լուծում են հանքային աղերը։
Արմատային մազիկները նույնպես մեխանիկական դեր են խաղում՝ նրանք ծառայում են որպես արմատի վերին մասի հենարան, որն անցնում է հողի մասնիկների արանքով։
Մանրադիտակի տակ, կլանման գոտում գտնվող արմատի խաչմերուկի վրա, նրա կառուցվածքը տեսանելի է բջջային և հյուսվածքների մակարդակը. Արմատի մակերեսին ռիզոդերմն է, ներքեւում՝ կեղևը։ Կեղևի արտաքին շերտը էկզոդերմա է, ներսից՝ հիմնական պարենխիման։ Նրա բարակ պատերով կենդանի բջիջները կատարում են պահեստավորման ֆունկցիա, սննդարար լուծույթներ են անցկացնում ճառագայթային ուղղությամբ՝ ներծծող հյուսվածքից մինչև փայտի անոթներ: Նրանք նաև սինթեզում են բույսի համար կենսական նշանակություն ունեցող մի շարք օրգանական նյութեր։ Կեղեւի ներքին շերտը էնդոդերմն է։ Էնդոդերմի բջիջներով կեղևից կենտրոնական գլան եկող սննդարար լուծույթներն անցնում են միայն բջիջների պրոտոպլաստով։

Կեղևը շրջապատում է արմատի կենտրոնական գլան: Այն սահմանակից է բջիջների շերտին, որոնք երկար ժամանակ պահպանում են բաժանվելու ունակությունը։ Սա պերիցիկլն է: Պերիցիկլային բջիջները առաջացնում են կողային արմատներ, ադնեքսային բողբոջներ և միջնակարգ կրթական հյուսվածքներ: Պերիցիկլից դեպի ներս՝ արմատի կենտրոնում, հաղորդիչ հյուսվածքներ են՝ բաստ և փայտ։ Նրանք միասին կազմում են ճառագայթային հաղորդիչ ճառագայթ:
Արմատի հաղորդիչ համակարգը ջուրն ու հանքանյութերը փոխանցում է արմատից դեպի ցողուն (վերընթաց հոսանք), իսկ օրգանական նյութերը՝ ցողունից դեպի արմատ (ներքև հոսանք)։ Այն բաղկացած է անոթային թելքավոր կապոցներից։ Փաթեթի հիմնական բաղադրիչներն են ֆլոեմի հատվածները (որոնցով նյութերը շարժվում են դեպի արմատ) և քսիլեմը (որով նյութերը շարժվում են արմատից)։ Ֆլոեմի հիմնական հաղորդիչ տարրերն են մաղի խողովակները, քսիլեմները՝ շնչափողերը (անոթները) և տրախեյդները։
Արմատային կյանքի գործընթացները
Ջրային տրանսպորտ արմատից
Արմատային մազերով ջրի կլանումը հողի սննդանյութերի լուծույթից և դրա անցկացումը շառավղային ուղղությամբ առաջնային ծառի կեղևի բջիջների երկայնքով էնդոդերմի անցման բջիջներով դեպի ճառագայթային անոթային կապոցի քսիլեմ: Արմատային մազերի կողմից ջրի կլանման ինտենսիվությունը կոչվում է ներծծող ուժ (S), այն հավասար է օսմոտիկ (P) և տուրգոր (T) ճնշման տարբերությանը. S=P-T։
Երբ օսմոտիկ ճնշումը հավասար է տուրգորային ճնշմանը (P=T), ապա S=0, ջուրը դադարում է հոսել դեպի արմատային մազի բջիջ։ Եթե հողի սննդարար լուծույթում նյութերի կոնցենտրացիան ավելի բարձր է, քան բջջի ներսում, ապա ջուրը կլքի բջիջները և տեղի կունենա պլազմոլիզ՝ բույսերը կթառամեն: Այս երեւույթը նկատվում է չոր հողի, ինչպես նաև հանքային պարարտանյութերի ավելորդ կիրառման պայմաններում։ Արմատային բջիջների ներսում արմատի ներծծող ուժը մեծանում է ռիզոդերմից դեպի կենտրոնական գլան, ուստի ջուրը շարժվում է կոնցենտրացիայի գրադիենտի երկայնքով (այսինքն՝ ավելի բարձր կոնցենտրացիան ունեցող վայրից ավելի ցածր կոնցենտրացիան ունեցող վայր) և ստեղծում է արմատային ճնշում։ որը բարձրացնում է ջրի սյունը քսիլոմային անոթների երկայնքով՝ առաջացնելով դեպի վեր հոսանք։ Այն կարելի է գտնել գարնանային տերևավոր կոճղերի վրա, երբ հավաքվում է «հյութը», կամ կտրված կոճղերի վրա։ Փայտից, թարմ կոճղերից, տերևներից ջրի արտահոսքը կոչվում է բույսերի «լաց»։ Երբ տերևները ծաղկում են, նրանք նաև ծծող ուժ են ստեղծում և ջուր են քաշում դեպի իրենց՝ յուրաքանչյուր անոթում գոյանում է ջրի շարունակական սյուն՝ մազանոթային լարվածություն։ Արմատային ճնշումը ջրի հոսանքի ստորին շարժիչն է, իսկ տերևների ներծծող ուժը վերինն է։ Դուք կարող եք դա հաստատել պարզ փորձերի օգնությամբ։
Ջրի կլանումը արմատներով
Թիրախ:պարզել արմատի հիմնական գործառույթը.
Ինչ մենք ենք անում:թաց թեփի վրա աճեցված բույս, թափահարեք նրա արմատային համակարգը և արմատները իջեցրեք մի բաժակ ջրի մեջ: Ջրի վրա բարակ շերտ լցնել՝ գոլորշիացումից պաշտպանելու համար։ բուսական յուղև նշեք մակարդակը:

Այն, ինչ մենք դիտարկում ենք.մեկ-երկու օր հետո բաքի ջուրն ընկավ նշագծից ցածր:

Արդյունք:ուստի արմատները ջուրը ներծծեցին ու հասցրին մինչեւ տերեւները։
Եվս մեկ փորձ կարելի է անել՝ ապացուցելով սնուցիչների արմատներով կլանումը։
Ինչ մենք ենք անում:բույսի ցողունը կտրում ենք՝ թողնելով 2-3 սմ բարձրությամբ կոճղ, կոճղի վրա դնում ենք 3 սմ երկարությամբ ռետինե խողովակ, իսկ վերևի ծայրին՝ 20-25 սմ բարձրությամբ կոր ապակե խողովակ։
Այն, ինչ մենք դիտարկում ենք.ապակե խողովակի ջուրը բարձրանում և դուրս է հոսում։
Արդյունք:սա ապացուցում է, որ արմատը հողից ջուր է ներծծում ցողունի մեջ:

Արդյո՞ք ջրի ջերմաստիճանը ազդում է արմատից ջրի կլանման արագության վրա:
Թիրախ:պարզել, թե ինչպես է ջերմաստիճանը ազդում արմատների աշխատանքի վրա:
Ինչ մենք ենք անում:մի բաժակը պետք է լինի տաք ջրով (+17-18ºС), իսկ մյուսը սառը ջրով (+1-2ºС):
Այն, ինչ մենք դիտարկում ենք.առաջին դեպքում ջուրն առատ է բաց թողնվում, երկրորդում՝ քիչ, կամ ամբողջովին դադարում է։
Արդյունք:սա ապացույց է, որ ջերմաստիճանը ուժեղ ազդեցություն ունի արմատների աշխատանքի վրա:

Տաք ջուրն ակտիվորեն ներծծվում է արմատներով։ Արմատային ճնշումը բարձրանում է:

Սառը ջուրը վատ է ներծծվում արմատներով: Այս դեպքում արմատային ճնշումը նվազում է:
հանքային սնուցում
Հանքանյութերի ֆիզիոլոգիական դերը շատ մեծ է։ Դրանք հիմք են հանդիսանում սինթեզի համար օրգանական միացություններ, ինչպես նաև կոլոիդների ֆիզիկական վիճակը փոխող գործոնները, այսինքն. ուղղակիորեն ազդում է պրոտոպլաստի նյութափոխանակության և կառուցվածքի վրա. հանդես գալ որպես կենսաքիմիական ռեակցիաների կատալիզատորներ; ազդում է բջջի տուրգորի և պրոտոպլազմայի թափանցելիության վրա. բույսերի օրգանիզմների էլեկտրական և ռադիոակտիվ երևույթների կենտրոններն են։
Հաստատվել է, որ բույսերի բնականոն զարգացումը հնարավոր է միայն սննդարար լուծույթում երեք ոչ մետաղների առկայության դեպքում՝ ազոտ, ֆոսֆոր և ծծումբ, և չորս մետաղներ՝ կալիում, մագնեզիում, կալցիում և երկաթ: Այս տարրերից յուրաքանչյուրն ունի անհատական արժեք և չի կարող փոխարինվել մյուսով: Սրանք մակրոէլեմենտներ են, դրանց կոնցենտրացիան բույսում 10 -2 -10% է: Բույսերի բնականոն զարգացման համար անհրաժեշտ են միկրոտարրեր, որոնց կոնցենտրացիան բջջում 10 -5 -10 -3% է: Դրանք են՝ բորը, կոբալտը, պղինձը, ցինկը, մանգանը, մոլիբդենը և այլն։ Այս բոլոր տարրերը գտնվում են հողում, բայց երբեմն՝ անբավարար քանակությամբ։ Այդ պատճառով հողի վրա կիրառվում են հանքային և օրգանական պարարտանյութեր։
Բույսը աճում և զարգանում է նորմալ, եթե արմատներին շրջապատող միջավայրը պարունակում է բոլոր անհրաժեշտ սննդանյութերը։ Հողը նման միջավայր է բույսերի մեծամասնության համար:
Արմատային շունչ
Բույսի բնականոն աճի և զարգացման համար անհրաժեշտ է, որ թարմ օդը մտնի արմատ: Եկեք ստուգենք, արդյոք դա այդպես է:
Թիրախ:Արդյո՞ք արմատները օդի կարիք ունեն:
Ինչ մենք ենք անում:Վերցնենք ջրով երկու միանման անոթ։ Յուրաքանչյուր անոթի մեջ տեղադրում ենք զարգացող սածիլներ։ Անոթներից մեկի ջուրն ամեն օր օդով ենք հագեցնում՝ օգտագործելով լակի ատրճանակ։ Երկրորդ անոթի ջրի մակերեսին լցնել բարակ շերտ բուսական յուղ, քանի որ այն դանդաղեցնում է օդի հոսքը ջրի մեջ։

Այն, ինչ մենք դիտարկում ենք.որոշ ժամանակ անց երկրորդ անոթի բույսը կդադարի աճել, թառամել և ի վերջո կմահանա:

Արդյունք:բույսի մահը տեղի է ունենում արմատի շնչառության համար անհրաժեշտ օդի բացակայության պատճառով:
Արմատային փոփոխություններ
Որոշ բույսերում պահուստային սննդանյութերը կուտակվում են արմատներում: Նրանք կուտակում են ածխաջրեր, հանքային աղեր, վիտամիններ և այլ նյութեր։ Նման արմատները ուժեղ են աճում հաստությամբ և ձեռք են բերում անսովոր տեսք։ Արմատային մշակաբույսերի առաջացմանը մասնակցում են և՛ արմատը, և՛ ցողունը։
Արմատներ
Եթե պահուստային նյութեր կուտակվում են հիմնական ընձյուղի հիմնական արմատում և ցողունի հիմքում, առաջանում են արմատային մշակաբույսեր (գազար)։ Արմատ առաջացնող բույսերը հիմնականում երկամյա են։ Կյանքի առաջին տարում նրանք չեն ծաղկում և շատ սնուցիչներ են կուտակում արմատային մշակաբույսերում։ Երկրորդին նրանք արագ ծաղկում են՝ օգտագործելով կուտակված սննդանյութերը և ձևավորում պտուղներ ու սերմեր։
արմատային պալարներ

Դալիում պահուստային նյութերը կուտակվում են պատահական արմատներում՝ առաջացնելով արմատային պալարներ։
բակտերիալ հանգույցներ
Երեքնուկի, լուպինի, առվույտի կողային արմատները յուրօրինակ փոփոխված են։ Բակտերիաները նստում են երիտասարդ կողային արմատներում, ինչը նպաստում է հողի օդից գազային ազոտի կլանմանը։ Նման արմատները ունենում են հանգույցների ձև: Այս բակտերիաների շնորհիվ այս բույսերը կարողանում են ապրել ազոտով աղքատ հողերի վրա և դրանք ավելի բերրի դարձնել։
ցցված

Միջմակընթացային գոտում աճող թեքահարթակը զարգացնում է ցցված արմատներ: Ջրից բարձր՝ նրանք անկայուն ցեխոտ հողի վրա մեծ տերևավոր կադրեր են պահում։
Օդ

Արևադարձային բույսերը, որոնք ապրում են ծառերի ճյուղերի վրա, զարգացնում են օդային արմատներ: Նրանք հաճախ հանդիպում են խոլորձների, բրոմելիադների և որոշ պտերերի մեջ։ Օդային արմատները ազատորեն կախված են օդում՝ չհասնելով գետնին և կլանելով խոնավությունը անձրևից կամ ցողից, որը թափվում է դրանց վրա։
Հետադարձիչներ

Սմբուկավոր և սոխուկավոր բույսերում, օրինակ՝ կրոկուսներում, բազմաթիվ թելիկ արմատների մեջ կան մի քանի ավելի հաստ, այսպես կոչված, հետ քաշվող արմատներ։ Կրճատելով՝ նման արմատները կորիզն ավելի խորն են քաշում հողի մեջ։
Սյունաձև

Ficus-ը զարգացնում է սյունաձև վերգետնյա արմատներ կամ օժանդակ արմատներ:
Հողը որպես արմատների բնակավայր
Բույսերի համար հողը այն միջավայրն է, որտեղից այն ստանում է ջուր և սննդանյութեր: Հողի մեջ օգտակար հանածոների քանակը կախված է մայր ապարի առանձնահատկություններից, օրգանիզմների ակտիվությունից, բուն բույսերի կենսագործունեությունից և հողի տեսակից։
Հողի մասնիկները մրցում են արմատների հետ խոնավության համար՝ պահելով այն իրենց մակերեսին։ Սա, այսպես կոչված, կապված ջուրն է, որը բաժանված է հիգրոսկոպիկ և ֆիլմի: Այն պահվում է մոլեկուլային ձգողականության ուժերի կողմից: Բույսին հասանելի խոնավությունը ներկայացված է մազանոթ ջրով, որը կենտրոնացած է հողի փոքր ծակոտիներում։
Հակառակ հարաբերություններ են զարգանում հողի խոնավության և օդային փուլի միջև։ Որքան մեծ են հողի ծակոտիները, այնքան լավ է այդ հողերի գազային ռեժիմը, այնքան քիչ խոնավություն է պահպանում հողը: Առավել բարենպաստ ջրային-օդային ռեժիմը պահպանվում է կառուցվածքային հողերում, որտեղ ջուրը և օդը գտնվում են միաժամանակ և չեն խանգարում միմյանց. ջուրը լցնում է կառուցվածքային ագրեգատների ներսում մազանոթները, իսկ օդը լցնում է նրանց միջև եղած մեծ ծակոտիները:
Բույսի և հողի փոխազդեցության բնույթը մեծապես կապված է հողի ներծծող կարողության հետ՝ քիմիական միացությունները պահելու կամ կապելու ունակության հետ:
Հողի միկրոֆլորան օրգանական նյութերը քայքայում է ավելի շատ պարզ կապեր, մասնակցում է հողի կառուցվածքի ձեւավորմանը։ Այս գործընթացների բնույթը կախված է հողի տեսակից, բույսերի մնացորդների քիմիական բաղադրությունից, միկրոօրգանիզմների ֆիզիոլոգիական հատկություններից և այլ գործոններից։ Հողի կառուցվածքի ձևավորմանը մասնակցում են հողի կենդանիները՝ անելիդները, միջատների թրթուրները և այլն։
Կենսաբանական և քիմիական գործընթացներՀողի մեջ ձևավորվում է օրգանական նյութերի բարդ համալիր, որը համակցված է «հումուս» տերմինով։
Ջրի կուլտուրայի մեթոդ
Ինչ աղեր են անհրաժեշտ բույսին և ինչ ազդեցություն ունեն դրանք նրա աճի և զարգացման վրա, պարզվել է ջրային կուլտուրաների հետ փորձի արդյունքում: Ջրային կուլտուրայի մեթոդը բույսերի մշակումն է ոչ թե հողում, այլ հանքային աղերի ջրային լուծույթում։ Կախված փորձի նպատակից՝ կարող եք լուծույթից բացառել առանձին աղ, նվազեցնել կամ ավելացնել դրա պարունակությունը։ Պարզվել է, որ ազոտ պարունակող պարարտանյութերը նպաստում են բույսերի աճին, ֆոսֆոր պարունակող պարարտանյութերը՝ մրգերի ամենավաղ հասունացումը, իսկ կալիում պարունակողները՝ տերևներից արմատներ օրգանական նյութերի ամենաարագ արտահոսքը: Այս առումով ազոտ պարունակող պարարտանյութերը խորհուրդ է տրվում կիրառել ցանքից առաջ կամ ամռան առաջին կեսին, ֆոսֆոր և կալիում պարունակող պարարտանյութեր՝ ամռան երկրորդ կեսին։
Օգտագործելով ջրային կուլտուրաների մեթոդը՝ հնարավոր եղավ պարզել ոչ միայն բույսի մակրոտարրերի կարիքը, այլև պարզել տարբեր միկրոտարրերի դերը։
Ներկայումս կան դեպքեր, երբ բույսերը աճեցվում են հիդրոպոնիկայի և աերոպոնիկայի մեթոդներով։
Հիդրոպոնիկան բույսերի մշակումն է խճաքարով լցված ամանների մեջ։ Սննդային լուծույթ պարունակող անհրաժեշտ տարրեր, ներքևից սնվում է անոթների մեջ։
Աերոպոնիկան բույսերի օդային մշակույթն է: Այս մեթոդով արմատային համակարգը օդում է և ինքնաբերաբար (մեկ ժամվա ընթացքում մի քանի անգամ) ցողվում է սննդանյութերի թույլ լուծույթով։
ՕՐԳԱՆՈԳՐԱՖԻԱ
Օրգանոգրաֆիա - կենդանի օրգանիզմի օրգանների նկարագրությունը.
Օրգան- մարմնի մի հատված, որը տարբերակված է մորֆոլոգիապես և մասնագիտացված ֆունկցիոնալ առումով. Երկու հիմնական գործառույթներին համապատասխան՝ տվյալ բույսի կյանքը պահպանելը և հետագա մի շարք սերունդներում վերարտադրելը, առանձնանում են վեգետատիվ և գեներացնող (վերարտադրողական) օրգանները։
Նախ կուսումնասիրենք վեգետատիվ օրգանները, որոնք կազմում են ընձյուղային համակարգը և արմատային համակարգը։ Արմատային համակարգի տարրը արմատն է։
Արմատ
Արմատը զարգացել է ավելի ուշ, քան կադրը և առաջացել է առաջին ցամաքային բույսերի՝ ռինոֆիտների ռիզոիդներից։
Արմատի տարբեր մասեր կատարում են տարբեր գործառույթներ և բնութագրվում են որոշակի մորֆոլոգիական առանձնահատկություններ. Այս մասերը կոչվում են գոտիներ: Երիտասարդ արմատի գոտիներ՝ բաժանման գոտի, աճի կամ ձգման գոտի, կլանման կամ կլանման և հաղորդման գոտի:
Դրսում գագաթային մերիստեմը ծածկված է արմատային գլխարկով։ Կափարիչի կենդանի պարենխիմային բջիջները լորձ են արտադրում: Լորձաթաղանթային նյութը բարձր հիդրատացված պեկտինային պոլիսախարիդ է: Այն կուտակվում է դիկտյոսոմների վեզիկուլներում, որոնք, միաձուլվելով պլազմային թաղանթին, ազատում են այն պլազմային թաղանթի և բջջաթաղանթի միջև ընկած տարածության մեջ։ Այնուհետև լորձը դուրս է գալիս բջջաթաղանթի մակերես, որտեղ ձևավորում է փոքրիկ կաթիլներ։
Այս լորձաթաղանթի շնորհիվ հողի մասնիկները կպչում են արմատի ծայրին և արմատի մազերին: Լորձը ծառայում է որպես ներծծող մակերես իոնների փոխանակման համար: Լորձաթաղանթը պաշտպանում է արմատի ծայրը վնասակար նյութերհողը և չորանալուց բարենպաստ պայմաններ է ստեղծում օգտակար բակտերիաների նստեցման համար։
Երբ արմատը աճում է և առաջ է մղում արմատի գլխարկը, գլխարկի ծայրամասի բջիջները մաքրվում են: Բջիջների կյանքի տևողությունը՝ արտաքին տեսքից մինչև շերտազատում, 4-9 օր է՝ կախված գլխարկի երկարությունից և բույսի տեսակից։
Արմատային գլխարկը վերահսկում է արմատների աշխարհընկալումը: Կափարիչի մեջ գրավիտացիայի ընկալման վայրը բջիջների կենտրոնական սյունն է՝ կոլումելլան, որում ամիլոպլաստները գործում են ստատոլիտների (ձգողականության ընկալիչների) դերում։
Գագաթային մերիստեմը բաղկացած է փոքր (10–20 մկմ տրամագծով) բազմաեզրային բջիջներից՝ խիտ ցիտոպլազմով և մեծ միջուկներով։
Սպորային բույսերն ունեն արմատային գագաթի մի սկզբնական տեսակ։
Սերմացու բույսերի արմատներում նկարագրված են գագաթային կազմակերպման երկու հիմնական տեսակ. Դրանցից մեկում բոլոր տարածքներն ունեն ընդհանուր սկզբնատառեր։ Սա գագաթային մերիստեմի բաց տեսակ է, որը հավանաբար ֆիլոգենետիկորեն պարզունակ է։ Երկրորդ տեսակի մեջ երեք մասերից յուրաքանչյուրը, այսինքն. կենտրոնական (կամ հաղորդիչ) մխոցը, առաջնային կեղևը և արմատի գլխարկն ունեն իրենց սկզբնատառերը: Սա գագաթային մերիստեմի փակ տեսակ է։
Առաջին փուլերում բջիջների աճի կազմակերպման համար պատասխանատու սկզբնատառերը հիմնականում կորցնում են իրենց միտոտիկ ակտիվությունը արմատների աճի հաջորդ փուլերում: Փոխարենը, բջիջները, որոնք որոշ չափով ավելի խորն են գտնվում արմատային մարմնում, ցուցադրում են միտոտիկ ակտիվություն: Այս դիտարկումները հիմք են տվել գագաթային մերիստեմում հանգստի կենտրոնի տեսության ձևավորմանը։ Համաձայն այս տեսության՝ արմատի մարմնի ամենադիստալ բջիջները (օրգանի ամենահեռու հատվածը՝ կենտրոնական կետից. չափը և ունեն ՆԱ և սպիտակուցի սինթեզի ցածր արագություն: Հանգստի կենտրոնն ունի կիսագնդաձև կամ սկավառակաձև ձև, ծավալով տատանվում է՝ կախված արմատի մեծությունից։ Հանգստի կենտրոնի բջիջները մոտավորապես 10 անգամ ավելի դանդաղ են բաժանվում, քան հարևան բջիջները:
Փորձնականորեն վնասված արմատներում քնած կենտրոնն ունակ է վերականգնել մերիստեմը։ Այսինքն՝ հանգստի կենտրոնը էական դեր է խաղում կառուցվածքի ձևավորման և արմատի զարգացման գործում։
Արմատի ծայրի առանձնահատկությունը գագաթային մերիստեմից առաջացող բջիջների երկայնական շարքերի տեղադրումն է։
Արմատի գագաթային մերիստամի շերտերը.
Դերմատոգեն(հուն. derma - մաշկ, genos - ծնունդ) - գագաթային մերիստեմի արտաքին շերտը, տարբերվում է ռիզոդերմի: Ճիշտ ձևի դերմատոգեն բջիջները.
Պլերոմա(հունարեն pleroma - լցնում) - գագաթային մերիստամի կենտրոնական մասը, որը տարբերվում է կենտրոնական գլանների բջիջների: Բջիջները երկարացվում են առանցքի երկայնքով:
արատավոր(հուն. periblema - ծածկույթ) - գագաթային մերիստեմի շերտ դերմատոգենի և պլերոմայի միջև, որը տարբերվում է առաջնային ծառի կեղևի բջիջների:
Առաջնային ծառի կեղևը տարբերվում է երկարացման գոտու մակարդակով։ Գաղութացման գոտում առաջնային կեղևը բաժին է ընկնում լայնակի հատվածի մեծ մասը: Այն կոչվում է «առաջնային»՝ առաջնային մերիստեմից ծագման պատճառով, «կեղև»՝ ծայրամասային դիրքի պատճառով։
Առաջնային ծառի կեղևի արտաքին բջիջները, որոնք գտնվում են ռիզոդերմի տակ, կոչվում են էկզոդերմա: Առաջնային ծառի կեղևի ներքին շերտը էնդոդերմա է:
Առաջնային ծառի կեղևի հիմնական մասը մեզոդերմա է: Մեզոդերմը ձևավորվում է պարենխիմային բջիջներով, որոնք գտնվում են թույլ տեղակայմամբ, ինչը նպաստում է արմատային բջիջների օդափոխմանը:
Գիմնոսպերմների և դիկոտրուկների մեծ մասի արմատները վաղ են թափում իրենց առաջնային կեղևը: Դրանցում առաջնային ծառի կեղեւի բջիջները մնում են պարենշիմային։ Ի հակադրություն, մոնոտիտների առաջնային ծառի կեղևը պահպանվում է արմատի ողջ կյանքի ընթացքում, և նրա բջիջները ձևավորում են երկրորդական լորձաթաղանթներ։
Արմատի կենտրոնը զբաղեցնում է առանցքային գլան՝ ստել։ Սկսում է տարբերակել աճի գոտում: Առանցքային մխոցի ձևավորումը սկսվում է դրա արտաքին շերտի ձևավորմամբ. pericycle.Պերիցիկլը պատկանում է առաջնային կողային մերիստեմներին, նրա բջիջները երկար ժամանակ պահպանում են մերիստեմատիկ ակտիվությունը: Սովորաբար պերիցիկլը ներկայացված է բջիջների մեկ շերտով, թեև այն կարող է լինել նաև բազմաշերտ:
Սերմաբույսերի մեծ մասում կողային արմատները դրված են պերցիկլում, առաջանում են ադնեքսային բողբոջների ռուդիմենտներ: Երկրորդային աճով տեսակների մեջ պերցիկլը մասնակցում է կամբիումի ձևավորմանը և առաջացնում է ֆելոգենի առաջին շերտը։
Պերիցիկլի տակ դրված են պրոկամբիալ բջիջներ, որոնցից առաջանում է առաջնային ֆլոեմը, հետագայում՝ առաջնային քսիլեմը։ Քսիլեմի ճառագայթների թիվը տարբեր տեսակների մեջ տատանվում է երկու, երեք, չորսից մինչև մի քանիսը: Ըստ քսիլոմային ճառագայթների քանակի՝ արմատները կոչվում են դիարխ, տրիարխ, տետրարխ, պոլիարխ։ Տետրարխի արմատները համարվում են առաջնային։ Մոնոկոտիկներն ունեն պոլիարխի կոթող:
Այսպիսով, ընդհանուր պլանԱրմատի առաջնային կառուցվածքը և հյուսվածքների մորֆոլոգիական տարբերակումը ցույց են տալիս արմատի մասնագիտացման և հարմարվողականության բարձր մակարդակ ջրի և հանքանյութերի կլանման և անցկացման գործառույթներին:
Կողմնակի արմատների ձևավորում
Կողային արմատները առաջանում են կենտրոնական գլանի ծայրամասում՝ գագաթային մերիստեմից տարբեր հեռավորությունների վրա։ Որովհետեւ դրանք դրված են արմատի խորը տեղակայված հյուսվածքներում, դրանց ծագումը կոչվում է էնդոգեն (հունարենից «ներսից ծնված»): Գիմնոսպերմների և անգիոսպերմների կողային արմատները դրված են պերցիկլում։ Պերիցիկլի բջիջները, որոնք ներգրավված են կողային արմատների ձևավորման մեջ, արմատային կամարն են:
Արմատային պրիմորդիայի բջիջների մի քանի շերտեր կարող են ձևավորվել նաև էնդոդերմայից։
Դիարխիկ արմատի մեջ կողային արմատը դրված է թմբուկի և քսակի միջև, եռիշխանական, քառակուսի արմատի մեջ՝ քսիլեմին հակառակ, միաշերտավորների բազմարխիկ արմատում՝ ֆլոեմին հակառակ։ Պերիցիկլային բջիջները բաժանվում են տարբեր ուղղություններով: Արդյունքում ձևավորվում է փոքր արմատային տուբերկուլյոզ: Չափի մեծանալով՝ արմատային ռուդիմենտը ճնշում է էնդոդերմի վրա։ Բայց էնդոդերմի պատյանները կոշտ են։ Հետեւաբար, էնդոդերմը դուրս է գալիս: Էնդոդերմի ուռուցիկությունը կոչվում է արմատային գրպան։
Արմատի պրիմորդիան անցնում է առաջնային ծառի կեղևով՝ արտազատելով ֆերմենտներ, որոնք ճանապարհին ոչնչացնում են կեղևի բջիջները։ Զարգացման վաղ փուլերում այս ռուդիմենտը կազմում է արմատի գլխարկը, գագաթային և առաջնային մերիստեմները: Ժամանակի ընթացքում պարենխիմային բջիջները քսիլեմի և ֆլոեմի տարրերի տարբերակելու պատճառով կողային և մայր արմատների կենտրոնական գլանները փոխկապակցված են:
Միակոթուններում առաջնային արմատային կառուցվածքը պահպանվում է ողջ կյանքի ընթացքում և երկրորդական հյուսվածքներ չեն առաջանում։ Տարիքի հետ բջիջների չափերը կարող են մեծանալ, բջջային թաղանթները խտանալ, սակայն արմատային կառուցվածքի ընդհանուր «պլանը» մնում է անփոփոխ։
Gymnosperms- ում և dicots- ում տեղի են ունենում երկրորդական փոփոխություններ, և ձևավորվում է երկրորդական արմատային կառուցվածք: Արմատի երկրորդական կառուցվածքի ձևավորումը կապված է հիմնականում կամբիումի գործունեության հետ։ Կամբիումը ձևավորվում է այն պրո-կամբիալ բջիջների բաժանումից, որոնք մնում են չտարբերակված առաջնային ֆլոեմի և առաջնային քսիլեմի միջև։ Այնուհետև պերիցիկլի բջիջները, որոնք տեղայնացված են քսիլոմային ելուստներից դուրս, նույնպես դրսևորում են կամբիալ ակտիվություն։
Կամբիումը երկրորդական քսիլեմային բջիջներն է դնում դեպի կենտրոն, իսկ երկրորդական ֆլոեմային բջիջները՝ ծայրամասում։ Այսպես են ձևավորվում հաղորդիչ կապոցները։ Դրանք ձևավորվում են այնքան, որքան առաջնային քսիլեմի ճառագայթները։ Աճող երկրորդական հյուսվածքների ճնշման տակ առաջնային ֆլոեմի բջիջները հարթվում են։ Առաջնային քսիլեմը պահպանում է իր կենտրոնական դիրքը և տեսանելի է որպես աստղանիշ: Ժամանակի ընթացքում առաջնային քսիլեմը նույնպես ջնջվում է։
Արմատի երկրորդական կառուցվածքում առաջնային հյուսվածքները չեն գործում որպես հաղորդիչ տարրեր։ Երկու ուղղություններով նյութերի հոսանքն իրականացվում է երկրորդական հյուսվածքների միջոցով։
Կամբիումը, որը սկիզբ է առնում pericycle-ից, ձևավորում է լայն պարենխիմալ ճառագայթներ, որոնք գտնվում են բաց գրավի կապոցների միջև: Սրանք հիմնական միջուկային ճառագայթներն են: Նրանք ապահովում են ֆիզիոլոգիական կապ արմատի կենտրոնական մասի և առաջնային կեղևի միջև։ Հետագայում կարող են ձևավորվել ավելի նեղ երկրորդական մեդուլյար ճառագայթներ, որոնք միացնում են երկրորդական քսիլեմն ու ֆլոեմը։
Երկրորդական փոփոխությունները տեղի են ունենում ոչ միայն կենտրոնական մխոցում: Հաղորդիչ հյուսվածքների հաստության ավելացման պատճառով առաջնային ծառի կեղևը դուրս է մղվում, պատռվում և թափվում էնդոդերմի հետ միասին։
Պերիցիկլի արտաքին մասում ֆելոգենը առաջանում և նստեցնում է ֆելլեման դեպի դուրս, իսկ բազմաշերտ ֆելոդերմը դեպի ներս։ Այսպիսով, երկրորդական կառուցվածք ունեցող դիկոտնիկների արմատները ծածկված են պերիդերմով։
Բազմամյա արմատներում ֆելոգենը հայտնվում է արմատի խորքում: Այնուհետեւ արմատը, ինչպես ցողունը, կազմում է ռիթիդոմ:
Երկրորդական արմատների աճի կարևոր էկոլոգիական ասպեկտնույն տեսակի տարբեր բույսերի արմատների բնական աճի ունակությունն է: Այնտեղ, որտեղ արմատները շփվում են միմյանց հետ, նրանք միավորվում են երկրորդական աճի միջոցով: Երբ արմատները միասին աճում են, հաստատվում է հաղորդիչ համակարգի շարունակականությունը երկու միաձուլված գործընկերների միջև: Արմատների միաձուլման շնորհիվ շատ ծառեր փոխկապակցված են, և եթե դրանցից մի քանիսը հատվում են, ապա կոճղերը երկար ժամանակ կենդանի են մնում։ Արմատների միաձուլումը նպաստում է վարակի արագ փոխանցմանը մի ծառից մյուսը։
Արմատների տարբերակումը արմատային համակարգերումդրսևորվում է տարբեր ձևերով. Առավել կարևոր է տարբերակումը հիմնական, կողային և պատահական արմատների:
Հատուկ խմբում առանձնանում են արմատները, որոնք առաջանում են բարձրագույն սպոր բույսերի գագաթային մերիստեմներում։ Ենթադրվում է, որ դրանք արմատների ամենաարխայիկ խումբն են և ամենամեծ նմանությունը պահպանել են առաջնային արմատների հետ, որոնք վերգետնյա և ստորգետնյա առանցքների մեկ ճյուղավորված համակարգի մաս էին կազմում։ Այս խմբի մեջ ընդգրկված արմատները կոչվում են ապիոգեն:
Ելնելով էվոլյուցիայի գործընթացում արմատային համակարգերի առաջացման հաջորդականությունից և մեծ համակարգային խմբերին պատկանող՝ մենք կարող ենք նախանշել երեքը. արմատային համակարգերի էվոլյուցիոն մակարդակը.
I. Բարձրագույն սպորների արմատային համակարգերը ներկայացված են բացառապես ապիգեն արմատային համակարգերով, որոնք հաջորդաբար փոխարինում են միմյանց, ինչը արմատային համակարգերում նորացման հիմնական մեխանիզմն է: Ճյուղավորումը գագաթային կամ կողային է, և հնարավոր է, որ ճյուղավորումների երկու տեսակներն էլ առաջացել են միմյանցից անկախ։ Նեղ մասնագիտացումով մետամորֆոզներ ու արմատներ չկան։
P. Գիմնոսպերմերի արմատային համակարգն առանձնանում է հիմնական արմատի առկայությամբ, որը կարող է կապված լինել սերմի և սաղմնային արմատով երկբևեռ սաղմի առաջացման հետ։ Նորացման մեխանիզմում ընդգրկված էին ոչ միայն ենթակա կարգերի կողային արմատները, այլև ռիզոգեն ադվենցիցիոն արմատները (այսինքն՝ արմատների վրա պատահական արմատները, ընձյուղների վրա պատահական արմատները մարմնամարզության մեջ նկատելի զարգացում չստացան): Մասնագիտացված և փոխակերպված արմատները հազվադեպ են:
Շ. Անգիոսպերմերի արմատային համակարգը բնութագրվում է ակամա արմատների, ինչպես նաև նեղ մասնագիտացում ունեցող մետամորֆոզների և արմատների ուժեղ զարգացմամբ։ Թարմացման մեխանիզմները բազմազան են և արդյունավետ:
Արմատային համակարգերի մետամորֆոզներ
Արմատային համակարգերի մետամորֆոզը անատոմիական և ձևաբանական առանձնահատկությունների մի շարք է, որոնք ձևավորվել են ֆիլոգենետիկորեն և ժառանգաբար ամրագրված և առաջացած ֆիզիոլոգիական գործառույթների փոփոխությամբ: Մետամորֆոզը (մետամորֆոզ, հունարեն - փոխակերպում) գենետիկ մակարդակում տեղի ունեցող փոփոխությունների արդյունք է, որը հանգեցնում է օրգանի, օրգան համակարգի կամ օրգանիզմի ձևաբանական և ֆունկցիոնալ վերակազմավորմանը:
I. Մետամորֆոզներ՝ կապված ատիպիկ ֆունկցիաների կատարման հետ.
1) շնչառական արմատները.
2) էպիֆիտների օդային արմատները.
3) ցցված արմատներ.
4) պահեստային արմատներ.
P. Մետամորֆոզներ՝ կապված հիպերֆունկցիայի հետ.
1) արմատային մշակաբույսեր.
2) արմատային կոներ;
3) արմատային հանգույցներ;
4) միկորիզա.
Արմատային կուլտուրաները բնորոշ են երկամյա երկշիկավոր բույսերին։ Սա ժամանակավոր անուն է։ Նրանք զարգանում են վեգետատիվ օրգաններից և ոչ մի կապ չունեն մրգերի հետ։ Արմատային բերքի ձևավորմանը մասնակցում են ցողունի ստորին հատվածը և արմատը։
Կյանքի առաջին տարում բույսը բուսականանում է, առաջանում է արմատային բերք։ Երկրորդ տարում ձևավորվում են ծաղիկներ, պտուղներ, սերմեր։ Սա նշանակում է, որ արմատային մշակաբույսը նյութերի մատակարարման օրգան է, որը թույլ է տալիս բույսին փոխանցել քնած շրջանը և ամբողջական զարգացումը հաջորդ տարվա համար։
Արմատների տեսակները.
1.Մոնոկամբիալ.
Սկզբնական արմատը դիարխի առանցքային գլան է: Արմատային բերքի ձևավորումը սկսվում է երկրորդական փոփոխություններով, որոնք կապված են մեկ կամբիալ օղակի սկզբնավորման և գործարկման և հետագա պարենխիմատիզացիայի հետ:
Մենք գիտենք, որ կամբիումը դնում է երկրորդական քսիլեմը դեպի ներս, իսկ երկրորդական փեղկը դրսից: Անատոմիական տարբերությունները վերաբերում են պահեստային հյուսվածքի զարգացման վայրին: Եթե ֆլոեմում տեղի է ունենում պարենխիմայի ուժեղ զարգացում, ապա ձևավորվում է արմատային բերքի ֆլոեմային տեսակ՝ գազարի տեսակ։ Եթե xylem-ում - xylem տեսակը - շաղգամ տեսակը:
2.Արմատային մշակաբույսերի պոլիկամբիալ տեսակ.
Ձևավորվում է մեկ տիպիկ կամբիալ օղակ, մնացածը պերիցիկլիկ ծագման լրացուցիչ օղակներ են։ Դրանք կարող են լինել մինչև 25: Կամբիումի այս լրացուցիչ օղակները ձևավորում են բաց գրավական անոթային կապոցներ: Որքան հեռու է օղակը առաջին կամբիալ օղակից, այնքան ավելի շատ պահեստային պարենխիմա է առաջանում: Պոլիկամբիալ արմատային մշակաբույսի օրինակ է ճակնդեղը:
Միաձույլ և պոլիկամբիալ արմատային մշակաբույսերի մեջ առաջնային կեղևը վաղ է մահանում և շերտավորվում: Ծածկույթի հյուսվածքը պերիդերմն է։
Այսպիսով, բոլոր արմատային մշակաբույսերի ընդհանուր հատկանիշը հաղորդիչ հյուսվածքներով ներծծված մեծ քանակությամբ պահեստային պարենխիմայի առկայությունն է: Հաղորդիչ և պահեստային հյուսվածքների միջև սերտ կապը ձեռք է բերվում երկրորդական աճի տարբեր փոփոխությունների միջոցով:
Որոշ բույսերում պահեստային արմատները ձևավորվում են կողային և պատահական արմատներից՝ արմատային կոների տեսքով։ Արմատային կոները (լյուբկա) կամբիալ են, ունեն առաջնային կառուցվածք; դիկոտիկներ - կամբիալ, ունեն երկրորդական կառուցվածք: Օրինակ՝ ֆլոեմի տիպի դահլիայի արմատային կոներ:
Արմատային կոները ամբողջ մակերևույթի վրա ձևավորում են պատահական արմատներ, իսկ վերին մասում՝ պատահական բողբոջներ և, հետևաբար, վեգետատիվ բազմացման օրգաններ են։
Հայտնի է, որ բույսերի աճը առավել հաճախ սահմանափակվում է ֆիքսված ազոտի անբավարար մատակարարմամբ։ N 2 կայուն մոլեկուլի տեսքով ազոտը կազմում է մթնոլորտի 80%-ը։ Նախքան ֆիքսումը (N 2-ի կրճատումը մինչև ամոնիակ NH 3), այս մոլեկուլը պետք է տրոհվի: Ազոտի ֆիքսումն իրականացվում է որոշ ազատ ապրող բակտերիաների կողմից, որոնք սպառում են հողի օրգանական նյութերը: Որոշ բակտերիաներ նստում են բարձր բույսերի արմատներին։ Բույսը «գրավում» է բակտերիաները օրգանական արմատային սեկրեցներով։ Իսկ միկրոօրգանիզմներն իրենց հերթին բույսին ապահովում են ֆիքսված ազոտով։ Երկու օրգանիզմների այս փոխշահավետ կապը կոչվում է սիմբիոզ։ Ամենից հաճախ արմատները կապեր են կազմում Rhizobium սեռի ազոտը ամրացնող բակտերիաների հետ: Բակտերիաների հետ սիմբիոզի մեջ կարող են մտնել երկշաքիլ և միաշաքայլ բույսերի ավելի քան 60 ընտանիք։ Բույսերի յուրաքանչյուր տեսակ ունի իր սիմբիոտիկ Rhizobium-ը: Բույսի և բակտերիաների փոխադարձ ճանաչումն իրականացվում է հատուկ բակտերիային միացնելով հատուկ սպիտակուց՝ լեկտին, որը գտնվում է տրիխոբլաստների մակերեսին։ Բակտերիաների կողմից արտազատվող աճի հորմոնների ազդեցության տակ արմատային մազերը դեֆորմացվում են։ Բակտերիաները հայտնվում են այս ոլորված բջիջների ներսում: Բջիջների ներսում բակտերիաները բազմանում են՝ ձևավորելով վարակիչ թել՝ փակված լնդերի պատյանում։ Այս թելը խորը թափանցում է արմատի մեջ։ Բակտերիաների նման ներթափանցման արդյունքը արմատային մեզոդերմի բջիջների ուժեղ բազմացումն է, ինչը հանգեցնում է հանգույցների առաջացմանը։ Հանգույցի ամենաարտաքին հատվածը երկար ժամանակ մնում է մերիստեմատիկ գոտի, իսկ բակտերիաները տեղակայված են հանգույցի ներքին մասում։ Բակտերիալ հյուսվածքն ինքնին ներկայացված է փոքր բջիջներով՝ մածուցիկ ցիտոպլազմով և հսկայական քանակությամբ բակտերիաներով։ Բակտերիալ հյուսվածքը շրջապատված է անոթային կապոցներով, որոնք կապված են արմատի կենտրոնական գլանին։ Նրանք. կա բակտերիաների և բույսերի միջև սննդանյութերի փոխանակման տրանսպորտային համակարգ: Ազոտ ամրացնող բակտերիաները բարենպաստ ազդեցություն ունեն բույսերի ազոտային սնուցման և հողի բերրիության վրա:
Արմատի ֆիզիոլոգիական ակտիվությունը, որը կապված է բույսին ջրով և հանքանյութերով ապահովելու հետ, կարող է ուժեղացվել հատուկ սնկերի հետ նրա սիմբիոտիկ կապով: Արմատների և սնկերի նման սերտ փոխշահավետ սիմբիոզ է միկորիզա.Միկորիզը հայտնի է անոթային բույսերի մեծ մասում: Միայն մի քանի ծաղկող ընտանիքներ չեն կազմում այն կամ հազվադեպ են առաջանում, օրինակ՝ կաղամբ, ցախ։
Միկորիզային սնկերը առանցքային դեր են խաղում անոթային բույսերի հանքային սնուցման մեջ: Միկորիզայի զգալի դերը ֆոսֆորի կլանման և փոխանցման գործում:
Երկկողմանի փոխադրման ժամանակ (ֆոսֆատները սնկերից բույսեր տեղափոխելը և ածխաջրերը հակառակ ուղղությամբ) կորուստը և կլանումը տեղի է ունենում բույսի և սնկերի պլազմային թաղանթների միջև ընկած ապոպլաստիկ գոտում:
Միկորիզային սնկերն օգտագործում են P-ի աղբյուրներ, որոնք սովորաբար հասանելի չեն բույսերին: Միկորիզը նպաստում է նաև բույսերի կողմից Zn, Mn, Cu-ի կլանմանը։ Հողի այս տարրերը թույլ շարժունակ են, և, հետևաբար, դրանցից քայքայված գոտիները արագ ձևավորվում են արմատային մազերի շուրջ: Միկորիզային սնկերի հիֆերի ցանցը տարածվում է յուրաքանչյուր արմատից մի քանի սանտիմետր և դրանով իսկ մեծացնում է արմատների ներծծող մակերեսը։
Կան միկորիզայի հետևյալ տեսակները.
1) էկտոմիկորիզա. բորբոսը հաստ ծածկով ծածկում է արմատի ամբողջ ծայրը և թափանցում միջբջջային տարածություններ.
2) էնդոմիկորիզա. բորբոսը ներխուժում է բջիջներ:
Էկտոմիկորիզը բնորոշ է ծառերի և թփերի առանձին ընտանիքներին (սոճին, ուռենին, հաճարենին): Էկտոմիկորիզան ձևավորվում է երիտասարդ, դեռևս չսուբերիզացված արմատների վրա: Էկտոմիկորիզային արմատները կարճ են, ճյուղավորված և ուռած տեսք ունեն։
Նման արմատներում արմատային մազիկների զարգացումը ճնշվում է, և գագաթային մերիստամի և արմատային գլխարկի ծավալը կարող է կրճատվել: Արմատային մազերի ֆունկցիան կատարվում է հիֆերով։
Էնդոմիկորիզային արմատները իրենց ձևով նման են սովորական արմատներին, բայց դրանցից տարբերվում են ավելի մուգ գույնով։ Սնկային հիֆերը թափանցում են արմատային ծառի կեղևի բջիջներ, որտեղ ձևավորում են վեզիկուլներ (ուռուցքներ) և արբուսկուլներ (ծառի նման ճյուղեր)։ Հետեւաբար, էնդոմիկորիզան հաճախ կոչվում է վեզիկուլյար-արբուսուլյար: Որոշակի պայմաններում սնկային հիֆերը կարող են «մարսվել» բջիջների կողմից: Արմատների մազածածկույթը պահպանվում է։ Սա միկորիզայի ամենատարածված տեսակն է: Այն հանդիպում է բոլոր անոթավոր բույսերի մոտավորապես 80%-ի մոտ, մասնավորապես՝ խոտաբույսերի, ներառյալ հացահատիկային բույսերի մոտ. թփերի, ծառերի մոտ (խնձորի ծառեր, թխկիներ ...): Երիտասարդ բարդի բույսի մեջ, օրինակ, ձևավորվում է վեզիկուլյար-արբոսկուլյար էնդոմիկորիզա, և երբ այն ծերանում է, զարգանում է պատյան էկտոմիկորիզա։
Heather Ericaceae-ին բնորոշ է էրիկոիդ-արբուտոիդ միկորիզը: Բորբոսը արմատի շուրջ ձևավորում է հիֆերի հզոր կուտակում, իսկ կողային բարակ հիֆերը ներթափանցում են արմատային կեղևի բջիջներ։ Այս միկորիզային սնկերը մեծապես նվազեցնում են հողի թունավորությունը շրթունքների համար՝ թույլ տալով այս բույսերին տեղավորվել թթվային, սննդանյութերով աղքատ հողերում:
Կա ևս մեկ տեսակ՝ խոլորձի էնդոմիկորիզա։ Բնության մեջ խոլորձի սերմերը բողբոջում են միայն համապատասխան սնկերի առկայության դեպքում։
Ընդհանուր առմամբ, բնական պայմաններում, միկորիզային ասոցիացիաները կանոն են, քան բացառություն:
Ամենահին անոթային բույսերի մնացորդների ուսումնասիրությունը ցույց է տվել, որ նրանց մոտ էնդոմիկորիզա տեղի է ունեցել ոչ պակաս հաճախ, քան ժամանակակից բույսերում: Սա թույլ տվեց ենթադրել, որ էվոլյուցիայի գործընթացում միկորիզայի առաջացումը կարող է լինել այն գործոններից մեկը, որը թույլ է տվել բույսերին գաղութացնել հողը: Հաշվի առնելով այս գաղութացման ընթացքում հողի վատ զարգացումը, կարելի է ենթադրել, որ բույսերի հանքային սնուցման հիմքում ընկած են միկորիզային սնկերը։ Միկորիզա ունեցող ժամանակակից պիոներ բույսերը աղքատ հողերում գոյատևելու ավելի մեծ հնարավորություններ ունեն:
Սնուցման միկոտրոֆ տեսակի բույսերը ավելի դիմացկուն են տեխնածին միջավայրում ծանր մետաղների առկայության նկատմամբ։
Մի քանի խոսք արևադարձային անտառային բույսերի արմատների մասին. Ավելի տարածված է, քան թեքված, տախտակի նման կրող արմատները: Արմատները հիշեցնում են տախտակներ, որոնք իրենց վերին ծայրերով հենված են ծառին։ Թեև ցցված արմատները պատահական արմատներ են, որոնք առաջանում են ցողունի վրա և աճում դեպի ներքև, տախտակի արմատների ձևավորումը կապված է կողային արմատների հետ, որոնք աճում են անմիջապես հողի մակերեսի տակ: Կողմնակի արմատները զարգացնում են ուղղահայաց ելքեր՝ լայնական կտրվածքով, որոնք հարում են ծառի բնին։
Տախտակի արմատների փայտը հաճախ ավելի կոշտ է, քան նույն ծառի մնացած փայտը, և հաճախ դրանք ծածկված են կեղևի բարակ շերտով: Դատելով այս արմատների խաչմերուկում տեսանելի աճի օղակներից՝ սկզբում դրանք նորմալ, կլորացված արմատներ են, բայց հետո վերին կողմում առաջանում են շատ ուժեղ խտացում։ Ծառի վրա տախտակի արմատների թիվը տատանվում է 1-ից 10-ի, բայց շատ դեպքերում դրանք երեք կամ ավելի են: Արևադարձային շրջաններում տախտակների արմատների առաջացումը հատկապես նկատելի է ճահճոտ անտառներում և վատ ցամաքեցված հողերում աճող ծառերի վրա: Հաճախ տախտակի արմատները օգտագործվում են արդյունաբերական նպատակներով: Այսպիսով, Ինդոնեզիայում աճող Koompassia excelsa ծառի տախտակի արմատները օգտագործվում են ճաշասեղաններ պատրաստելու համար:
ՓԱԽՍՏԸ
Ծիլը, ինչպես արմատը, բույսի հիմնական վեգետատիվ օրգանն է։ Կրակոցը կատարում է օդային սնուցման ֆունկցիա, կարող է կատարել մի շարք այլ գործառույթներ և ունակ է փոխակերպման։
Վեգետատիվ չփոփոխված կադրը բաղկացած է առանցքային մասից՝ ցողունից; տերեւները եւ բողբոջները: Հիմնական արտաքին հատկանիշը, որը տարբերում է ընձյուղը արմատից, ընձյուղի տերեւավորությունն է։
Ցողունի այն հատվածը, որը կապված է կողային օրգանների (ճյուղեր, տերևներ և այլն) առաջացման հետ. հանգույց.Ցողունի հատվածը հարակից հանգույցների միջև միջհանգույցներ.Ներքին անկյուն տերևի և ցողունի միջև տերևի առանցք.Ծիլերի յուրաքանչյուր կրկնվող տարր (տերևով հանգույց և բողբոջ տերևի առանցքում և միջհանգույց) ձևավորվում է. մետամեր.Այսպիսով, կրակոցը բաղկացած է մի շարք մետամերներից, այսինքն. ունի մետամերիկ կառուցվածք։
20-րդ դարի 90-ականների սկզբին հանգույց և միջհանգույց հասկացությունների փոխարեն ամերիկացի գիտնականներն առաջարկեցին. ցողունային տարր.Ցողունային տարրը ցողունի հատվածն է տերևային պրիմորդիայի եզրերի միջև։ Բացահայտվել է կրակոցի գագաթնակետում, որտեղ դեռ միջանցք չկա: Նրանք. «ցողունային տարր» նոր հասկացությունը համապատասխանում է «մետամեր» հասկացությանը։
Ծիլերի գագաթային մերիստեմի կառուցվածքն ավելի բարդ է, քան արմատինը։ Իրոք, ծիլում գագաթային մերիստեմը ոչ միայն ձևավորում է առաջնային հյուսվածքների բջիջները, այլև մասնակցում է տերևային պրիմորդիայի և առանցքային բողբոջների ձևավորմանը, որոնք այնուհետև վերածվում են կողային ընձյուղների: Այս մերիստեմը պաշտպանված չէ արմատային գլխարկի նման կառուցվածքով:
Գագաթների տեսակները նույնն են, ինչ արմատում է՝ միասկզբ, զոնալ, աստիճանավոր։ Շերտերի թիվն ավելի մեծ է, քան արմատում:
Էվոլյուցիոն պրիմիտիվ մի սկզբնական գագաթ: Այս տեսակի գագաթնակետի օրինակը հանդիպում է ձիու պոչերում: Մեկ մեծ երկարաձգված գագաթային բջիջը խիստ վակուոլացված է: Այն բաժանվում է իր մակերեսներին զուգահեռ ուղղություններով (բացառությամբ ամենաարտաքին մակերեսի)։ Մոտակա ածանցյալ բջիջները նույնպես բարձր վակուոլացված են, բայց երբ նրանք բաժանվում են, հայտնվում են փոքր բջիջներ՝ խիտ պրոտոպլաստներով։ Նման բջիջները հայտնաբերվում են ցողունի գագաթի ծայրամասի երկայնքով, որտեղ դրված են տերևային պրիմորդիաները։
Ծաղկավոր բույսերի մեծ մասի ընձյուղի վեգետատիվ գագաթը կառուցված է ինչպես տունիկա՝ մարմին՝ երկու գոտի, որոնք տարբերվում են բջիջների բաժանման հարթություններում: Տունիկա(լատ. - պատյան) ներառում է բջիջների արտաքին շերտերը, որոնք բաժանվում են անտիկլինալ (մակերեսին ուղղահայաց) և ապահովում մակերեսի բարձրացում։ Շրջանակ(լատ. - մարմին) - թունիկի տակ գտնվող բջիջների կենտրոնական խումբը, որը բաժանվում է տարբեր հարթություններում, կազմելով նկարահանման հիմնական ծավալը: Թունիկայի մարմինը և յուրաքանչյուր շերտ ունի իր սկզբնատառերը։ Տունիկայում սկզբնատառերը տեղադրվում են կենտրոնում՝ առանցքի երկայնքով։ Երկշերտ տունիկա ունեն հիմնականում երկշերտ տունիկա, միաշերտները՝ միաշերտ, ավելի քիչ հաճախ՝ երկշերտ։
Թեև թիկնոց-կեղևի գաղափարը նպաստում է բուն գագաթային մերիստեմում բջիջների տեղակայման և աճի մասին մեր ըմբռնմանը, հարակից տարածքներում տարբեր բջջային գոտիների ճանաչումն օգնում է բացահայտել մերիստեմի ներդրումը հյուսվածքների և օրգանների տարբերակման գործում: կրակոցը. Բջջաբանական տարբեր բնութագրերով գոտիների գագաթային մերիստեմում առկա է ցիտոհիստոլոգիական գոտիավորում.
Կորպուսը համապատասխանում է բջիջների մի շրջանի, որը կոչվում է կենտրոնական մայր բջիջների գոտի.Այս գոտու բջիջները բարձր վակուոլացված են, ինչը կապված է միտոտիկ ակտիվության համեմատաբար ցածր արագության հետ: Կենտրոնական մայրական բջիջների գոտին շրջապատված է ծայրամասային մերիստեմով, որը մասամբ առաջանում է թունիկից, մասամբ կորպուսից։ Պրոտոդերմային հիստոգենը առաջանում է թունիկայի արտաքին շերտից։ Ծայրամասային մերիստեմը առաջանում է գագաթային սկզբնատառերի կողային ածանցյալներից և կենտրոնական մայր բջիջներից։ Միջուկային մերիստեմը ձևավորվում է կենտրոնական մայր բջիջների ծայրամասի երկայնքով բաժանումների արդյունքում, որը կոչվում է շերտ. անցումային գոտի.Ծայրամասային գոտու բջիջներն ունեն խիտ պրոտոպլաստներ, հետևաբար՝ ունեն բարձր միտոտիկ ակտիվություն։
Հատկապես բարձր ակտիվությամբ որոշ վայրերում դրվում են տերևային պալարներ (կամ առաջնային պալարներ):
Նոր տերևային պրիմորդիայի առաջացումը կապված է բջիջների բաժանման ուղղության փոփոխության հետ, որին նախորդում է ցիտոպլազմայի ծայրամասային շերտերում միկրոխողովակների կողմնորոշման փոփոխությունը։ Օսմոտիկ ճնշումը նույնպես ազդում է պրիմորդիայի աճի վրա։
Ակտիվ աճի շրջանում ընձյուղի գագաթային մերիստեմն այնքան արագ է ձևավորում տերևային պրիմորդիա, որ հանգույցներն ու միջհանգույցները սկզբում չեն տարբերվում։ Աստիճանաբար տերևների շերտերի միջև ընկած հատվածները սկսում են աճել, ձգվել և միջհանգույցների ձև ստանալ։ Միևնույն ժամանակ, տերևների ամրացման տարածքները ձևավորվում են հանգույցներով:
Ներքևում, որտեղ հայտնվում են տերևների ռուդիմենտները, նկատվում է վակուոլացում։ Սա ցույց է տալիս առաջնային ծառի կեղևի և կորիզի տարբերակման սկիզբը: Ուժեղ վակուոլիզացիան առաջնային ծառի կեղևում և բշտիկում ցույց է տալիս մերիստեմատիկ գոտու առկայությունը, որը կոչվում է մնացորդային մերիստեմ: Կարծես թե դա գագաթային մերիստեմի մնացորդ է, որը պահպանվել է ավելի տարբերակված հյուսվածքների մեջ: Մնացորդային մերիստեմում տերևային ռուդիմենտի տեղադրման վայրից ներքև, երկայնական բաժանումների միջոցով, առանց բջիջների հետագա աճի լայնությամբ, ձևավորվում են առաջին պրոկամբիումի փոքր-ինչ ձգված բջիջները։
Զարգացման հաջորդ փուլերում մնացորդային մերիստեմում ավելի ու ավելի շատ պրոկամբիալ թելեր են տարբերվում։ Ցողունի տվյալ մակարդակի բոլոր անոթային կապոցների ձևավորումից հետո մնացորդային մերիստեմը տարբերվում է միջֆասիկուլյար պարենխիմայի: Հանգույցներում մնացորդային մերիստամի մի մասը դառնում է տերևի ճեղքվածքային պարենխիմա:
Պրոկամբիումը կարող է դրվել շարունակական օղակով կամ առանձին թելերով: Եթե պրոկամբիումն ընկած է շարունակական օղակի մեջ, ա ցողունային կառուցվածքի առանց ճառագայթների (ոչ կապոցային):Եթե պրոկամբիումը դրված է առանձին թելերով, ա ճառագայթի տեսակը ցողունային կառուցվածքի.Հետևաբար, առաջնային հաղորդիչ հյուսվածքները գտնվում են ցողունում տարբեր ձևերով. առաջին դեպքում՝ շարունակական օղակում, երկրորդ դեպքում՝ առանձին հաղորդիչ կապոցներով, որոնք առանձնացված են հիմնական պարենխիմի հատվածներով՝ միջուկային ճառագայթներով։
Գրավի անոթային կապոցներում առաջնային փեղկը հայտնվում է պրոկամբիալ լարի արտաքին մասում, իսկ առաջնային քսիլեմը՝ ներքին մասում։ Ֆլոեմի հետագա տարբերակումը տեղի է ունենում կենտրոնաձիգ. թմբուկի նոր տարրերը հայտնվում են ավելի մոտ ցողունի կենտրոնին: Xylem-ը տարբերվում է հակառակ ուղղությամբ՝ կենտրոնախույս:
Ամբողջ ստելը կամ ցողունի կենտրոնական մասը բաղկացած է հաղորդիչ հյուսվածքներից, միջուկից (երբեմն այն փլուզվում է) և պերցիկլից։ Պերիցիկլից դուրս առաջնային կեղևն է: Առաջնային ծառի կեղևի կազմը (և այն ձևավորվում է ծայրամասային մերիստեմից) ներառում է քլորենխիմա, երբեմն սեկրետորային տարրեր։ Ծայրամասային մերիստեմի ամենաարտաքին շերտը չի կազմում էկզոդերմա, ինչպես արմատում, այլ տարբերվում է կոլենխիմայի, որը գտնվում է կամ թելերի տեսքով կամ շարունակական օղակի մեջ (կախված ցողունի կառուցվածքի տեսակից՝ կապակցված կամ առանց կապոցների): Առաջնային ծառի կեղևի ամենաներքին շերտը էնդոդերմն է: Բայց ցողունում այն կարգավորիչ ֆունկցիա չի կատարում, ինչպես արմատում։ Օսլայի հատիկները կուտակվում են էնդոդերմում, և այդ պատճառով այն կոչվում է օսլա պարունակող թաղանթ: Երբեմն այն պարունակում է կալցիումի օքսալատի բյուրեղներ:
Պերիցիկլը արագորեն կորցնում է իր մերիստեմատիկ ակտիվությունը և վերածվում սկլերենխիմայի, որը առաջանում է շարունակական շերտով կամ թելերով՝ կախված ցողունի կառուցվածքի տեսակից։
Միջուկը ներկայացված է բարակ պատերով պարենխիմալ բջիջներով, որոնցում պահուստային սնուցիչներ են պահվում։ Կան իդիոբլաստային բջիջներ՝ լցված բյուրեղներով, լորձով և այլն։ Միջուկի ծայրամասային մասը կոչվում է perimedullary գոտի(հունարեն peri - շուրջը, medulla - միջուկ): Պերիմեդուլյար գոտու բջիջներն ավելի փոքր են և երկար ժամանակ պահպանում են իրենց կենսագործունեությունը։
Միակոթուններում սկզբնական տեսակը ցողունային կառուցվածքի կապոցային տեսակն է, այսինքն. procambium-ը գտնվում է թելերի մեջ: Բայց երկրորդական մերիստեմը` կամբիումը, երբեք չի առաջանում, և, հետևաբար, երկրորդական հաղորդիչ հյուսվածքներ չկան: Նույնիսկ խտացման դեպքում ցողունը ձևավորվում է միայն առաջնային հյուսվածքներով: Միակոթանների հաստ ցողունը (արմավենիներ, հալվե) զարգանում է հիմնական պարենխիմի բջիջների բաժանման և ձգման շնորհիվ։ Այս աճը կոչվում է ցրված երկրորդական աճ:
Միաշերտավորների մոտ առաջնային ծառի կեղևը և կենտրոնական գլանն իրարից չեն բաժանվում հստակ սահմանով, ինչպես երկկոտորակներում, և ստելը բարձրանում է էպիդերմիսի տակ։
Միակոթունները բնութագրվում են ցողունային կառուցվածքի ցրված տեսակով, երբ փակ անոթային կապոցները ցրված են ցողունով մեկ։ Միջհանգույցների երկարացման պրոցեսում ձգվում և քայքայվում են ֆլոեմի և քսիլեմի առաջնային տարրերը (պրոտոֆլոեմ և պրոտոքսիլեմ), ինչի հետևանքով կապոցի քսիլեմային կողմում առաջանում է մեծ օդակեր խոռոչ։ Հետևաբար, լաբորատոր հետազոտություններում, երբ մենք դիտեցինք կապոցների տեսակները, եգիպտացորենի ցողունի խաչմերուկի մշտական պատրաստման վրա պարզ դարձավ, որ հասուն անոթային կապոցը ներառում է երկու լայն մետաքսիլեմային անոթ, օդային խոռոչ և մետաֆլոեմ: Յուրաքանչյուր կապոց պարփակված է սկլերենխիմային բջիջների երեսպատման մեջ:
Միակոթուններում ցողունի միջուկը հաճախ մեռնում է, և դրա տեղում առաջանում է խոռոչ։ Այդպիսի խոռոչ ցողունը լավ սահմանված հանգույցներով կոչվում է ծղոտե.Սոլոմինային բնորոշ է մեխանիկական հյուսվածքի ուժեղ զարգացումը, որը օղակ է կազմում ցողունի ծայրամասում։ Հասուն բույսերում քլորենխիման քայքայվում է, իսկ ծղոտը ձեռք է բերում ոսկեդեղին գույն՝ բնորոշ սկլերենխիմային։ Ընդհանրապես, մենակոթունները բնութագրվում են ցողունի ուժեղ սկլերեֆիկացմամբ, երբ բոլոր հյուսվածքների մինչև 30%-ը սկլերենխիմա են, և կարող է լինել կենդանի պարենխիմային բջիջների թաղանթների մասնակի կնճիռ:
Երկաթոռների ցողունում երկրորդական աճը կամբիումի գործունեության արդյունք է։ Կամբիումը հայտնվում է ընձյուղի աճի առաջին տարվա վերջում և նպաստում ցողունի խտացմանը։ Կամբիումը մասամբ առաջանում է անոթային կապոցներում գտնվող պրոկամբիումից և մասամբ միջֆասկուլյար պարենխիմայից։ Կախված առաջացման վայրից, կամբիումի հատվածները կոչվում են ֆասիկուլյար և միջֆասիկուլյար կամբիում։ Ապագա կամբիալ բջիջները միջֆասիկուլյար գոտիներում որոշվում են կամբիալ գործունեության մեկնարկից շատ առաջ։ Կամբիումը սկզբում ձևավորվում է ֆասիկուլուսին հարող բջիջներում, այնուհետև տարածվում է մեդուլյար ճառագայթի խորքում։ Արդյունքում միջֆասիկուլյար և ֆասիկուլյար կամբիումը ցողունում ձևավորում է շարունակական կամբիալ օղակ։
Երկրորդական աճի երեք տեսակ կա.
1) Aristolochia Chircason տեսակը.
Նախնականը ցողունային կառուցվածքի փնջի տեսակն է: Առաջնային անոթային կապոցները բաժանված են լայն միջուկային ճառագայթներով: Միավորված կամբիումը ձևավորում է երկրորդական հաղորդիչ հյուսվածքներ, իսկ միջֆասիկուլյար կամբիումը կազմում է ճառագայթային պարենխիման: Այս ճառագայթային կառուցվածքը պահպանվել է:
2) Հելիանտուս արևածաղկի տեսակ.
Ճառագայթային կառուցվածքի տեսակը նույնպես նախնականն է։ Բայց երկրորդական հաղորդիչ հյուսվածքները ձևավորվում են ինչպես ֆասիկուլյար, այնպես էլ միջֆասիկուլյար կամբիումով: Միջֆասկուլյար կամբիումի կողմից ձևավորված երկրորդական անոթային կապոցները տարբերվում են առաջնայիններից իրենց ավելի փոքր չափերով և կապոցի վերևում պերիցիկլային ծագման մեխանիկական հյուսվածքի բացակայությամբ: Կամբիումի երկարատև գործողությամբ, ավելի ու ավելի շատ են երկրորդական կապոցները: Նրանք աճում են, միաձուլվում միմյանց հետ՝ կազմելով երկրորդական հաղորդիչ հյուսվածքների շարունակական օղակ։
3) Տիլիա լորենի տեսակ.
Սկզբնական տեսակը ցողունային կառուցվածքի առանց ճառագայթների տեսակն է: Ընդհանուր կամբիալ օղակը ձևավորում է երկրորդական հաղորդիչ հյուսվածքներ: Սակայն ժամանակ առ ժամանակ կամբիումը ձևավորում է պարենխիմալ բջիջներ, որոնք էլ ավելի են տարբերվում երկրորդական ճառագայթների։
ցողունի կառուցվածքը բազմամյա բույսեր
Փայտային և թփուտային երկշիկավորների, ինչպես նաև փշատերևների մոտ երկրորդական խտացումը շարունակվում է երկար տարիներ, իսկ որոշ տեսակների մոտ կոճղերը հասնում են մի քանի մետրի տրամագծի։ Թանձրացումները կապված են երկարաժամկետ գործող կամբիումի և մասամբ ֆելոգենի՝ երկու երկրորդական կողային մերիստեմների գործունեության հետ: Բազմամյա փայտային բույսի ցողունում տեղագրական առումով կարելի է առանձնացնել երեք հիմնական մաս՝ կեղև, փայտ և բութ։ Կեղևի և փայտի միջև սահմանն անցնում է կամբիումի երկայնքով:
Կամբիումի բջիջները, ի տարբերություն բնորոշ մերիստեմատիկ բջիջների, միշտ բարձր վակուոլացված են: Կամբիալ բջիջների ցիտոպլազմայի վիճակը փոխվում է եղանակների հետ:
Կամբիալ գոտում կամ կամբիալ օղակում հայտնաբերվում են կամբիալ բջիջների երկու ձև՝ ուղղահայաց ուղղությամբ երկարացված սպինաձև բջիջներ (սկզբնագրեր) և հորիզոնական ուղղությամբ երկարացված գնդաձև բջիջներ՝ ճառագայթների սկզբնատառեր։ Եթե spindle բջիջները դասավորված են կանոնավոր հորիզոնական շարքերում, կամբիումը կոչվում է աստիճանավոր կամբիում: Եթե spindle- ի տեսքով բջիջների տեղադրման մեջ նման կարգ չկա, ապա կամբիումը ոչ շերտավոր է:
Longline cambium- ը առաջացնում է երկարաձիգ փայտ և երկարակյաց երկրորդային փխրուն: Անշերտ կամբիումը առաջացնում է ոչ շերտավոր փայտ և ոչ շերտավոր երկրորդական փեղկ: Էվոլյուցիոն առումով, երկարատև անտառները և փլոեմները համարվում են ավելի բարձր մասնագիտացված, քան ոչ հատակային փայտերը:
Ցողունի հաստացման հետ մեծանում է նաև կամբիումի շերտերի շրջագիծը։ Դա հնարավոր է դարձել ճառագայթային ուղղությամբ կամբիալ բջիջների բաժանման շնորհիվ։
Կամբիալ բջիջների շարքում միջին սկզբնական շերտը մեկուսացված է։ Բաժանման արդյունքում նրա բջիջները ձևավորում են ածանցյալներ ներքին, այնուհետև ներս արտաքին ուղղություն. Նույն ժամանակահատվածում 2-3 բջիջ նստում են դեպի ներս, և միայն մեկ բջիջ դրված է արտաքին ուղղությամբ։ Արդյունքում ավելի շատ փայտ է ձևավորվում, քան բաստը:
Ճառագայթային բջիջները (սկիզբները) կազմում են առանցքային պարենխիմալ ճառագայթներ՝ առաջնային, որոնք ձգվում են միջուկից մինչև կեղև; իսկ երկրորդական միջուկային ճառագայթներն ավելի կարճ են, քանի որ առաջանում են ավելի ուշ առաջացած ճառագայթային բջիջներից:
Հիմնական ճառագայթները կարող են լինել մի շարք և բազմաշար; միատարր (այսինքն՝ բաղկացած նույն բջիջներից) և տարասեռ (ճառագայթը ներառում է կառուցվածքով տարբեր բջիջներ)։
Հիմնական ճառագայթների իմաստը : այն սննդանյութերի (օսլա, լիպիդներ) պահեստավորման վայր է, որը դեր է խաղում քնած ժամանակաշրջանում (օրինակ՝ ձմռանը)։
Միջուկային ճառագայթներն ապահովում են ջրի և հանքային աղերի ճառագայթային փոխանցում քսիլեմից ապոպլաստի երկայնքով դեպի կամբիում և երկրորդական ֆլոեմ, և հակառակ ուղղությամբ՝ սննդանյութերի սիմպլաստի երկայնքով:
Եվ վերջապես գազափոխանակությունը տեղի է ունենում միջբջջային տարածություններով։
Փայտ
Փայտը ձևավորվում է կամբիումով և աճում է կենտրոնախույս ուղղությամբ: Որովհետեւ Մեր գոտում կամբիումը պարբերաբար աշխատում է. այն սկսում է գործել վաղ գարնանը և դադարում է գործունեությունը ամառվա վերջում, այնուհետև փայտի մեջ տեսանելի է համակենտրոն շերտավորումը: Գարնանը, աճող սեզոնի սկզբում, ձևավորվում է վաղ փայտանյութ: Նրա անոթները լայն են, բարակ պատերով, կարող են զգալի քանակությամբ ջուր կրել։ Ջուրն անհրաժեշտ է աճը սկսելու համար, հատկապես նոր բջիջների չափը մեծացնելու համար (օրինակ՝ զարգացող տերևային բջիջները): Ուշ փայտը պարունակում է ավելի քիչ անոթներ. Նրանք ավելի նեղ են և հաստ: Նույն տարվա վաղ փայտից ուշ փայտից անցումը կարող է աստիճանական լինել: Այնուամենայնիվ, ուշ փայտի սահմանը մեկ տարվա վաղ փայտի հետ հաջորդ տարիմիշտ սուր և հստակ տեսանելի խաչմերուկում: Արդյունքում ձեւավորվում է տարեկան աճ (տարեկան օղակ կամ տարեկան շերտ)։
Ըստ տարեկան (տարեկան) աճի մեջ անոթների տեղակայման՝ առանձնանում են փայտի հետևյալ տեսակները.
Եթե անոթները գտնվում են հիմնականում վաղ փայտի մեջ, իսկ ուշ փայտի մեջ դրանք բացակայում են կամ տեղադրվում են միայն փոքր անոթներ, օրինակ՝ կաղնու, հացենի, սա օղակաձեւ փայտ.
Եթե անոթները հավասարաչափ բաշխված են տարեկան աճի ընթացքում, չնայած դրանց տրամագիծը նվազում է ուշ փայտի մեջ, սա ցրված անոթային փայտ.Հայտնաբերվել է կեչի, թխկի, բարդիի մեջ։
Փայտի այս տեսակների միջև կան նաև անցումային ձևեր.
Քանի որ փայտը կատարում է հաղորդիչ, մեխանիկական և պահեստավորման գործառույթներ, այն նաև ներառում է, բացի քսիլեմային տարրերից, մեխանիկական հյուսվածք՝ սկլերենխիմայի տեսքով և պահեստային հյուսվածք՝ հիմնական պարենխիմայի տեսքով: Բոլոր անատոմիական տարրերի կառուցվածքը, դրանց թիվը և տեղաբաշխումը տարբեր բույսերի փայտին տալիս են որոշակի տաքսոնոմիկ խմբին բնորոշ առանձնահատկություններ:
Օրինակ, բույսերը, որոնք ավելի առաջադեմ են էվոլյուցիայի առումով, ունեն մասնագիտացված մեխանիկական հյուսվածք (sclerenchyma): Էվոլյուցիոն պարզունակ երկշաքիլավորները (մագնոլիաները) չունեն փայտի մանրաթելեր։ Մեխանիկական դերն այս դեպքում խաղում են տրախեիդները։
Որոշ բույսերի փայտի մեջ կան անցումային ձևեր տրախեյդներից դեպի լիբրֆորմ (libriform-ը փայտի մասնագիտացված մեխանիկական տարրեր են, որոնք բաղկացած են ծայրերում մատնանշված խիտ լորձաթաղանթներով պրոզենխիմային բջիջներից): Կա նաև cloisonne libriform։ Նրա սկզբնական spindle-ձև բջիջը հաստ ուղղահայաց թաղանթներով բաժանված է առանձին կարճ կենդանի բջիջների: Նման հյուսվածաբանական տարրերը մորֆոլոգիական և ֆունկցիոնալ առումով մոտ են փայտային պարենխիմային։
Փայտի պարենխիման կատարում է պահեստավորման և մասամբ հաղորդիչ դեր: Պարենխիմայի բջիջները կենդանի են: Փայտի պարենխիման տեղադրված է հորիզոնական և ուղղահայաց շարքերում: Հորիզոնական շարքերը կազմում են հիմնական ճառագայթները: Մենք արդեն դիտարկել ենք դրանց առանձնահատկությունները: Ուղղահայաց շարքերը ինքնին կազմում են փայտային պարենխիման:
Եթե փայտի պարենխիման ցրված է տարեկան աճի ողջ ընթացքում, սա ցրված պարենխիմա.Բնորոշ է լորենի, կաղնու, տանձի փայտի համար:
Եթե փայտի պարենխիման գտնվում է անոթների շուրջը, սա paratracheal parenchyma.Մոխիր, թխկի փայտ։
Տարեկան աճի արտաքին սահմանին է գտնվում տերմինալ պարենխիմա.Օրինակ՝ ուռենու, խեժի, մագնոլիայի փայտ։
Նրա կառուցվածքի առանձնահատկությունը արյան անոթների բացակայությունն է։ Շնչափողի տարրերը չծակված են և ներկայացված են տրախեիդներով, որոնք կատարում են ինչպես հաղորդիչ, այնպես էլ մեխանիկական դերեր:
Փշատերև տրախեիդները բնութագրվում են մեծ եզրագծով ծակոտիներով՝ տորուսով։ Տրախեիդները երկար են՝ սրածայր ծայրերով։ Վաղ տրախեիդները մեծ են, բարակ պատերով։ Հետագայում տրախեիդները շառավղային հարթեցված են, հաստ պատերով։
Մենք տեսնում ենք, որ մարմնամարզիկների և դիկոտնիկների փայտը կառուցվածքային զգալի տարբերություններ ունի։ Հետեւաբար, այն բաժանված է երկու հիմնական խմբի. Գիմնոսպերմերի փայտը դասակարգվում է որպես փափուկ, իսկ դիկոտնիկների փայտը՝ կոշտ: Այս «կոշտ» և «փափուկ» տերմինները պարտադիր չէ, որ սահմանափակվեն խտության և կարծրության աստիճանով: Տարբերությունները կապված են, ինչպես արդեն ասացինք, կառուցվածքային առանձնահատկությունների հետ։
Թեքված կամ կոր կոճղերի մեջ առաջանում է ռեակտիվ փայտ: Դրա ձևավորումը պայմանավորված է ճյուղերի և կոճղերի հակվածությամբ՝ դիմակայելու բեռներին, որոնք առաջանում են, երբ դրանք թեքված են, այսինքն. դրա ձևավորումը կապված է բույսի այս մասերի ուղղման գործընթացի հետ:
Ռեակտիվ փայտի զարգացման վրա ազդող ամենակարևոր գործոններն են գրավիտացիոն ուժերը և էնդոգեն աճի խթանիչների բաշխումը:
Ռեակտիվ փայտը սովորական փայտից տարբերվում է ինչպես անատոմիական, այնպես էլ քիմիապես: Փայտի ռեակտիվ բջիջներն ավելի մեծ չափով ենթարկվում են պատերի խտացման և խտացման: Փշատերևների մեջ այն ավելի խիտ և մուգ է, քան շրջապատող հյուսվածքը: Նրա տրախեիդներն ավելի կարճ են, քան սովորական փայտից:
Ցողունային փայտի ռադիոակտիվությունը նվազում է արտաքին շերտերից դեպի միջուկ: Փայտի աղտոտվածության որոշակի աճ կա բեռնախցիկի հիմքից մինչև վերև:
Կեղևում և կադրերում ռադիոակտիվ նյութերի բավականաչափ բարձր պարունակությունը վկայում է դրանց արտաքին օդային աղտոտվածության մասին:
Հաչել
Այս տերմինը ներառում է կամբիումից դուրս գտնվող բոլոր հյուսվածքները: Սա արդեն երկրորդական հաչոց է, քանի որ. ձևավորվում է երկրորդական մերիստեմով՝ կամբիումով (ի տարբերություն առաջնային, որը ձևավորվում է առաջնային մերիստեմի բջիջների տարբերակման ժամանակ)։
Երկրորդային ծառի կեղևի կազմը ներառում է երկրորդական թմբուկը, սկլերենխիման (բաստի մանրաթելեր և քարքարոտ բջիջներ) և հիմնական պարենխիման։ Մեխանիկական տարրերի բազմությունը կոչվում է կոշտ բամբակ,հաղորդիչ տարրեր և հիմնական պարենխիմա - փափուկ բշտիկ.
Երկրորդական ծառի կեղևում հայտնաբերվում են արտազատիչ բջիջներ և խեժային ջրանցքներ: Երկրորդային ծառի կեղևի հիմնական գործառույթը պլաստիկ նյութերի հաղորդումն է, ինչպես նաև պաշտպանիչ գործառույթը։
Երկրորդական փունջը կազմում է միջքաղաքային ծավալի շատ ավելի փոքր մասնաբաժին, քան երկրորդական քսիլեմը: Դա բացատրվում է նրանով, որ նախ կամբիումը քսիլեմից ավելի քիչ ֆլոեմ է ձևավորում, և երկրորդ՝ հին, չգործող ֆլոեմն աստիճանաբար ջախջախվում է։
Առանցքային օրգանի շրջագծի մեծացման պատճառով պարենխիմային ճառագայթները լայնանում են և նմանվում եռանկյունու, որի գագաթն ուղղված է դեպի կամբիումը։ Նման պարենխիման կոչվում է ընդլայնված (լատ. dilatatio - ընդլայնում): Պարենխիմայի այս դասավորությունը կանխում է բշտիկի պատռումը, քանի որ ցողունը հաստությամբ աճում է:
Փշատերևների մեջ երկրորդական փլեմն ավելի պարզ կառուցվածք ունի։ Գիմնոսպերմների բշտիկում հյուսվածքաբանական տարրերը հավասարաչափ բաշխված են և, իհարկե, առկա են խեժային ջրանցքներ։
Ֆելոգենից դուրս գտնվող բոլոր հյուսվածքները (նկատի ունի խցանափայտի ամենաներքին կամբիումը) դադարում են ջուր և հանքանյութեր ստանալ, քանի որ խցանի բջիջները դառնում են ենթաբերինիզացված: Այս բոլոր պերիդերմները կեղևի մնացորդներով (պարենխիմի և ֆլոեմի մահացած բջիջներ) կազմում են արտաքին (արտաքին) կեղևը։ Կեղևի կենդանի մասը, որը գտնվում է կամբիումի և ֆելոգենի ամենաներքին շերտի միջև, կոչվում է ներքին կեղև:
Այսպիսով, «Հյուսվածքներ» թեմայից մեր գիտելիքների հիման վրա մենք ուսումնասիրեցինք ցողունի անատոմիական կառուցվածքը՝ որպես կրակոցի առանցքային օրգան։
Այժմ դիտարկենք կրակոցի կողային օրգանի՝ տերևի կառուցվածքը:
ԹԵՐԹ
Տերեւը բարձրագույն բույսերի ֆոտոսինթետիկ հիմնական օրգանն է։ Տերևի կառուցվածքը և նրա գործառույթները սերտորեն փոխկապակցված են:
Ֆոտոսինթեզի ռեակցիաների ընդհանուր հավասարումից.
CO 2 + H 2 O chl-l → hv (CH 2 O) n + O 2
կարող ենք եզրակացնել, որ 1) տերևներին անհրաժեշտ է CO 2 և ջրի աղբյուր. 2) տերևները պետք է հարմարեցվեն կլանմանը արեւային էներգիանրանք պետք է ունենան քլորոֆիլ; 3) O 2-ը կթողարկվի որպես ռեակցիայի արտադրանքներից մեկը. 4) ածխաջրերը կամ պետք է պահվեն պահեստում կամ տեղափոխվեն գործարանի այլ մասեր: Ցուցակը շատ մասնագիտացված մարմին է, որը համապատասխանում է այս բոլոր պահանջներին:
Տերևի ամբողջ էվոլյուցիան, որպես վեգետատիվ օրգան, հետևեց լույսի լավագույն օգտագործման հարմարվողականության զարգացման ուղուն:
Ըստ էվոլյուցիայի միկրոֆիլային գծի՝ տերեւը համարվում է ռինոֆիտ տեսակի արխայիկ բույսերի տելոմայի առաջացում և պահպանում է առանցքային օրգանի կառուցվածքը։
Ըստ էվոլյուցիայի մակրոֆիլ գծի՝ տերեւն առաջացել է հարթեցման, տելոմների մեկ հարթության մեջ տեղադրելու և դրանց հետագա միաձուլման արդյունքում։ Միաժամանակ կորավ երկարաժամկետ գագաթային աճի և ճյուղավորվելու ունակությունը։ Որո՞նք են տերևի մորֆոլոգիական և անատոմիական առանձնահատկությունները, որոնք առաջացել են երկար էվոլյուցիայի արդյունքում:
Հասուն տերևը սովորաբար բաղկացած է տերևի շեղբից և բշտիկից: Ոտնաթաթան- տերևի ցողունային նեղ հատվածը տերևի շեղբի և ընձյուղի հանգույցի միջև, որի օգնությամբ տերևը կողմնորոշվում է տարածության մեջ և տեղադրվում է առավել բարենպաստ լույսի նկատմամբ: Ծաղկի միջով տերևի շեղբը և ցողունը միացված են։ Հաղորդող և մեխանիկական հյուսվածքները լավ զարգացած են կոթունում։ Տերեւաթափի ժամանակ բշտիկում առաջանում է բաժանարար շերտ։
Տերեւի ամենացածր մասը՝ ցողունով հոդավորված, կոչվում է թերթի հիմք:Տերևի հիմքը կարող է ունենալ տարբեր ձևեր: Ամենից հաճախ այն կարծես փոքր խտացում է և կոչվում է տերևի բարձիկ:Ավելի հաճախ թերթիկի հիմքում կան տարբեր ձևերև զուգակցված կողային ելքերի չափերը՝ ճարմանդներ։ Ստիպուլները կարող են լինել տերևաձև և ծառայել որպես լրացուցիչ յուրացնող մակերես։ Կտրուկները կարող են լինել ասեղաձև և հանդես գալ որպես բույսերի պաշտպանություն:
Առկա են թեփուկավոր, թաղանթավոր բշտիկներ։ Էվոլյուցիոն առումով նկատվում է բշտիկների կրճատում, իսկ էվոլյուցիոն առումով երիտասարդ բույսերում դրանք վաղ են ընկնում կամ իսպառ բացակայում են:
Որոշ հացահատիկներում՝ նեխուրի (հովանոցային), տերևի հիմքն աճում է և ձևավորում փակ կամ բաց խողովակ՝ տերևի պատյան։ Տերևի պատյանը նպաստում է միջքաղաքային մերիստամի երկարատև պահպանմանը և ծառայում է որպես լրացուցիչ հենարան ընձյուղի համար։
Տերևի հիմնական մասը տերևի շեղբն է։ Տերևի վերին և ստորին կողմերի անհավասար լուսավորության արդյունքում թիթեղն ունի թիկունքային կառուցվածք, այսինքն. Տերևի վերին կողմի կառուցվածքը՝ փորային, տարբերվում է ստորին կողմի կառուցվածքից՝ մեջքային։ Տերևի վերին կողմը կոչվում է փորային, քանի որ. երիկամի մեջ այս կողմը ներքին է և թեքված է դեպի կրակոցի առանցքը։ Բողբոջի մեջ գտնվող տերևի ստորին, մեջքային կողմը բույսից դեպի դուրս է շրջված: Այսպիսով, թիկունքային սիմետրիա ունեցող տերեւը երկկողմանի է կամ երկկողմանի կամ երկկողմանի: դիկոտիկներին բնորոշ.
Շառավղային համաչափություն ունեցող տերևը հավասարազոր է: Նման թանձրացած, գլանաձեւ տերեւները հանդիպում են Crassulaceae-ում։ Հացահատիկային, սոխին բնորոշ են միակողմանի կամ միակողմանի միակողմանի տերևները։
Հաշվի առեք տերևի շեղբի անատոմիական կառուցվածքը.
Ինչպես արմատը և ցողունը, տերևը բաղկացած է ամբողջական, հաղորդիչ և հիմնական հյուսվածքներից: Որովհետեւ տերևը սովորաբար չունի երկրորդական աճ (բացառությամբ կոթունների և խոշոր երակների մի փոքր աճի), այն պահպանում է էպիդերմիսը որպես ծածկույթի հյուսվածք:
Ստոմատները հիմնականում հանդիպում են տերևի ստորին մասում։ Երկկոտիլ տերևներում ստոմատները ցրված են տերևի ամբողջ մակերեսով առանց տեսանելի հերթականության։ Միաբնակ և փշատերև տերևներում դրանք բաշխված են տերևի երկայնական առանցքին զուգահեռ շարքերով։ Որոշ հացահատիկների էպիդերմիսում կան շարժիչ կամ շարժիչ բջիջներ։ Նրանք ավելի մեծ են, քան նորմալ էպիդերմիսի բջիջները: Վակուոլը զբաղեցնում է գրեթե ամբողջ բջիջը։ Տուրգորի կորստով այս բջիջները կծկվում են և նպաստում տերևի ծալմանը կամ ոլորմանը:
Տերևի սայրի հիմնական հյուսվածքի մեծ մասը գտնվում է մեզոֆիլում: Մեզոֆիլը տարբերակված է. Palisade mesophyll- ը գտնվում է ափսեի վերին մասում, սպունգային `մինչև ներքև: Նման կառուցվածքով տերևները երկդիմացական կամ թիկունքային են։
Եթե փալիզադային մեզոֆիլը գտնվում է տերևի երկու կողմերում, տերևը միակողմանի է կամ միակողմանի:
Հացահատիկային մշակաբույսերի մեզոֆիլում հստակ տարբերակում չկա փալիզադի և սպունգանման քլորենխիմայի: Մեզոֆիլային բջիջները շառավղով դասավորված են կապոցների շուրջ:
Հաղորդող տերևային կապոցները կոչվում են երակներ, իսկ երակների ճյուղավորվող համակարգը վենացիա.Անոթային կապոցների դասավորության առանձնահատկությունները տերևների շեղբերներում որոշում են օդափոխության տարբեր տեսակներ, որոնք կարող են լինել երկատված, զուգահեռ, կամարաձև, արմավենու և փետաձև:
Կախված նրանից, թե ինչպես են փոխկապակցված ավելի բարձր ճյուղավորվող կարգերի երակները, առանձնանում են բաց (երկբաժան) և փակ օդափոխություն։ Դիխոտոմային օդափոխությամբ երակները ճյուղավորվում են երկատված, հեռանում են սուր անկյան տակ և հասնում տերևի շեղբի եզրին՝ ոչ մի տեղ միաձուլվելով հարևան երակների հետ: Փակ օդափոխության դեպքում փոքր երակները անաստոմոզների (հատումների) արդյունքում կազմում են ցանց, որը ներթափանցում է ամբողջ ափսեը կամ դրա մեծ մասը։
Տերևները արմավենու և փետրավոր երակով, և դրանք հիմնականում երկշաքիլավոր են, ունեն միջին, ամենամեծ երակ: Երակը բաղկացած է առաջնային քսիլեմից և առաջնային փեղկից, որոնք առաջանում են պրոկամբիումից և միավորվում գրավի փաթեթների մեջ: Կամբիում հաճախ ձևավորվում է քսիլեմի և ֆլոեմի միջև, բայց այն չի գործում: Հետևաբար, քսիլեմը և ֆլոեմը առաջնային են:
Փաթեթները տերեւում ստեղծում են շարունակական համակարգ՝ կապված ցողունի հաղորդիչ համակարգի հետ։ Հետևաբար, քսիլեմը միշտ ուղղված է տերևի մորֆոլոգիապես վերին կողմին, մինչդեռ ֆլոեմը միշտ ուղղված է մորֆոլոգիապես ստորին կողմին:
Միջին հատվածը կողային միացված է ավելի փոքր կողային երակներով։ Նրանցից յուրաքանչյուրը կապված է նույնիսկ ավելի փոքրերի հետ, իսկ նրանք, իրենց հերթին, բաժանվում են նույնիսկ ավելի փոքրերի և այլն։ Ճյուղավորվող պատվերների թիվը դիկոտուտներում տատանվում է 2-ից 5 կամ ավելի: Ամենափոքր ճյուղերը կազմում են բջիջներ, որոնց ներսում փակ են փոքր տարածքներմեզոֆիլ - արեոլա.
Բաց երկատված օդափոխության համակարգում փակ բջիջներ չկան:
Մոնոկոտ տերևներում (զուգահեռ օդափոխությամբ) երկայնական երակները փոխկապակցված են ավելի փոքր երակներով՝ կոմիսուրալ կապոցներով, որոնք դասավորված են պարզ լայնակի կամուրջների տեսքով։ Փոքր երակներում հաղորդիչ հյուսվածքների թիվը աստիճանաբար նվազում է։ Փաթեթների ծայրերում քսիլեմային տարրերը հաճախ տարածվում են ավելի մեծ հեռավորության վրա, քան փլոեմները: Երակների ծայրերում գտնվող քսիլեմը սովորաբար բաղկացած է կարճ շնչափող տարրերից, ֆլոեմը՝ կարճ նեղ մաղի տարրերից և արբանյակային մեծ բջիջներից։
Մեզոֆիլում տեղայնացված փոքր անոթային կապոցները շրջապատված են կոմպակտ ծալված բջիջների մեկ կամ մի քանի շերտերով, որոնք կազմում են կապոցի պատյանը: Փաթեթի պատյանները կարող են լինել պարենխիմալ կամ սկլերենշիմային: Որոշ տեսակների մոտ նկատվում է ֆասիկուլային պատյանների բջջային պատերի սուբերինիզացիա, ինչը ենթադրում է, որ թաղանթի բջիջները կարող են գործել որպես էնդոդերմա:
Կոլենխիման և սկլերենխիման գտնվում են մեկ կամ երկու կողմի խոշոր երակներում: Մեծ երակների հետ կապված հյուսվածքը բարձրանում է տերևի մակերևույթից և ձևավորում ելուստներ։ Երակների ելուստների միջև պարփակված հյուսվածքի տարածքը կոչվում է «միջկողային գոտի» (լատ. ելուստի անվանումն է՝ «կոստա»՝ կողոսկր)։
Հաստ տերևներով տեսակների մեջ մեզոֆիլում հայտնաբերվում են տարբեր սկլերիդներ, որոնք գործում են որպես միջատներ։
Հացահատիկային տերևներին բնորոշ է սկլերենխիմի ուժեղ զարգացումը։
Gymnosperms- ի տերեւները կառուցվածքով ավելի քիչ բազմազան են, քան անգիոսպերմերի տերեւները: Մտածեք, օգտագործելով սոճու ասեղների օրինակը, թե ինչպես են ձմռանը ցածր ջերմաստիճանը հանդուրժելու հարմարվողականությունը և մի քանի տարի ապրելու ունակությունը արտացոլվում անատոմիական կառուցվածքում: Ասեղն ունի հաստ պատերով էպիդերմիս՝ հզոր կուտիկուլայով և խորը ընկղմված ստոմատներով: Որոշ փշատերևների (Cupressaceae) կողային բջիջներն ունեն կուտիկուլյար սրածայր, որը կախված է ստոմատայի պահակային բջիջների վրա։ Ստոմատները դասավորված են ասեղների բոլոր կողմերից ուղղահայաց շարքերով: Էպիդերմիսի տակ գտնվում են հիպոդերմիսի հաստ պատերով, մասամբ պատված բջջային թաղանթները: Մեզոֆիլը ծալված է: Այն պարունակում է խեժի անցումներ: Հաղորդող կապոցները գտնվում են ասեղի կենտրոնում: Սովորաբար երկու, հազվադեպ մեկ: Այս կապոցները միացված են սկլերենշիմային շերտով: Փաթեթները շրջապատված են տրախեիդներից և պարենխիմային բջիջներից բաղկացած փոխներարկման հյուսվածքով: Տրանսֆուզիոն հյուսվածքը (լատ. transfusio - փոխներարկում) կապված է հաղորդիչ կապոցների և մեզոֆիլի միջև ջրի և սննդանյութերի փոխադրման հետ։
Տրանսֆուզիոն հյուսվածքը շրջապատված է հաստ պատերով էնդոդերմայով։ Էնդոդերմը հաճախ ունենում է կասպարյան շերտեր զարգացման վաղ փուլերում և սուբերիկ թիթեղներ՝ հետագա փուլերում: Հասուն վիճակում էնդոդերմային բջիջներն ունեն երկրորդական lignified մեմբրաններ:
Տերևի շեղբի անատոմիական կառուցվածքը բացահայտում է զգալի պլաստիկություն: Կառուցվածքը փոխվում է՝ կախված բույսի գոյության պայմաններից և բույսի վրա տերևների տեղադրությունից։
Մենք արդեն նշել ենք ժամանակակից բուսաբանության զարգացման ուղղություններից մեկը՝ բույսերի էկոլոգիական անատոմիան և ասել, որ բջջի կենդանի պարունակությունն է՝ պրոտոպլաստը, որն առաջին հերթին արձագանքում է շրջակա միջավայրի պայմանների փոփոխություններին: Այսպիսով, հյուսիսային բույսերի բջիջները ավելի քիչ վակուոլացված են և ունեն մեծ քլորոպլաստներ։ Ստրոմայում կան բազմաթիվ օսլայի հատիկներ։ Քլորոպլաստների թիլաոիդ համակարգը ավելի քիչ զարգացած է, քան բարեխառն լայնությունների բույսերում։ Հյուսիսային բույսերում միտոքոնդրիաների թիվը 2-2,5 անգամ ավելի է։ Առավել զարգացած են EPR-ը և Golgi-ի ապարատը։ Հյուսիսային բույսերի մեզոֆիլի բջիջների ենթամանրադիտակային կառուցվածքի նման առանձնահատկությունները բնութագրվում են որպես կարևոր հարմարվողական հատկություններ, որոնք նպաստում են բույսերի գոյատևմանը կոշտ կլիմայական պայմաններում:
Խոնավության պակասին հարմարեցված բույսերում նկատվում են սառը, հաստ բջջային թաղանթներ, հատկապես էպիդերմիսում։ Խոշոր բջջային էպիդերմիսի խիստ հաստացած արտաքին թաղանթները զբաղեցնում են բջջային խոռոչի կեսը։ Էպիդերմիսի վերևում կուտիկուլային հաստ շերտ է: Մակերեւույթից կուտիկուլը ծածկված է մոմի հաստ թեփուկներով, որոնք որոշում են տերևների կապտավուն երանգը։
Ստոմատները ընկղմված են էպիդերմիսի մազիկներով արտամղված իջվածքների մեջ։
Սուկուլենտ բույսերն ունեն ջրի պահպանման հյուսվածք։
Հետաքրքիր հարմարեցումներ փափուկ քինոայի (Աֆրիկա) տերևներում խոնավությունը փակելու համար: Վերին և ստորին կողմերից տերևը ծածկված է բազմաշարք տեղադրված վեզիկուլյար մազիկների շարունակական շերտով։ Այս շերտի հաստությունը զգալիորեն գերազանցում է մեզոֆիլի հաստությունը և պաշտպանում է կենդանի հյուսվածքները չորանալուց։ Մազերը կարողանում են գիշերը պահել օդի խոնավությունը և բարելավել կոշտությունը ջրային ռեժիմըբույսեր.
Մեր փռված անտառային գոտու քինոայի տերևներն ու ցողունը ծածկված են բազմաթիվ պղպջակների նման մազիկներով՝ թողնելով փոշոտ ծածկույթի տպավորություն, բայց նրանք չեն կազմում այնպիսի խիտ բազմաշարք շերտ, ինչպես փափուկ քինոայում։
Կինոայի երկու տեսակների տերևների համեմատության օրինակով երևում է, որ նմանատիպ տաքսոնոմիկ միավորների համակարգային առանձնահատկությունները, երբ դրանք տեղավորվում են էկոլոգիական տարբեր պայմաններում, անհավասար զարգացում են ստանում և իրենց բնույթով հարմարվողական են։
Անտառային բույսերի (օրինակ՝ հրեշների) տերևներն ունեն ծակոտկեն տերևների շեղբեր և երկար կոթուններ։ Ավելորդ ջուրը միշտ հեշտությամբ հոսում է: Ստոմատները բաց են։ Palisade գործվածք մեծ օդային խոռոչներով:
Բացի ջերմաստիճանից և խոնավությունից, մեկ այլ կարևոր աբիոտիկ գործոն լույսն է, որին բույսն արձագանքում է կառուցվածքային փոփոխություններով։
Արևի ուղիղ ճառագայթների տակ զարգացող տերևներն ավելի փոքր են, քան ստվերային տերևները, բայց ավելի հաստ են, քան ստվերային տերևները՝ պալիսադային մեզոֆիլի ուժեղ զարգացման արդյունքում:
Այսպիսով, էվոլյուցիայի ընթացքում հաստատված շրջակա միջավայրի պայմաններին հարմարվողականությունը կապված է քանակական տարբերությունների հետ, բայց չի փոխում կազմակերպման սկզբունքները:
Տերևների օնտոգենեզ
Տերեւները դրված են փոքր պալարների տեսքով՝ որպես գագաթային մերիստեմի կողային ելքեր։ Տերեւային տուբերկուլյոզի առաջացմանը մասնակցում է ոչ միայն տունիկը, այլեւ մարմնի բջիջները։ Տերևային պրիմորդիան աճում է սկզբնական ելուստից դեպի վեր՝ կոնաձև կամ ասեղաձև ելքի տեսքով։ Շուտով, տերևների ռուդիմենտները շրջապատում են գագաթային մերիստեմը՝ պաշտպանելով այն ինչպես մեխանիկորեն, այնպես էլ շնչառության ընթացքում թողարկվող ջերմությունից:
Վաղ տերևների աճը սովորաբար բաժանվում է գագաթային և եզրային (մարգինալ): Առաջինը կապված է պրիմորդիայի երկարացման հետ, երկրորդը՝ կողային աճի հետ, որի արդյունքում առաջանում են ափսեի երկու մաս։ Ըստ այդմ, աճող պրիմորդիան ունի գագաթային մերիստեմ գագաթին և երկու եզրային մերիստեմ՝ առանցքի կողքերում։ Պրիմորդիայի գագաթային աճը երկար չէ։ Գագաթի և ափսեի եզրերի աճին հաջորդում է միջանկյալ աճը։
Բարդ տերևում եզրային մերիստամի ակտիվությունը տեղայնացված է առանձին կենտրոններում, որոնցից յուրաքանչյուրը կազմում է առանձին թերթիկ։
Տերեւաթիթեղի զարգացումն ավելի արագ է ընթանում, քան բշտիկի աճը, որը հետագայում առաջանում է միջանկյալ աճով։
Եթե միջքաղաքային և եզրային գոտիների հիմքի սահմանին առաջանում է կծկում, ապա առաջանում է կոթունի տերեւ։ Խոշոր կոթունի մեծ մասը ձևավորվում է միջքաղաքային գոտիով։ Եթե նման նեղացում (միջկալային և եզրային գոտիների հիմքի սահմանին) չի առաջանում, առաջանում է նստադիր տերեւ։
Միաշերտ բույսերի տերևային ռուդիմենտում ծայրամասային մերիստեմներ գրեթե չեն ձևավորվում, երկար ժամանակ բնորոշ է միջքաղաքային աճը։ Ուստի տերևի շեղբի հիմնական ձևը գծային է:
Թիթեղների տարբեր շերտերում բջիջների բաժանման և երկարացման արագության տարբերությունները հանգեցնում են բազմաթիվ միջբջջային տարածությունների և մեզոֆիլային բնորոշ կառուցվածքի ձևավորմանը:
Այլ հյուսվածքների զարգացումը փոխկապակցված է հաղորդիչ հյուսվածքների զարգացման հետ: Պրոկամբիումը միաժամանակ առանձնանում է տերևի և աճի կոնի հարակից տարածքում՝ ձևավորելով շարունակական թել։ Այն կազմում է առաջնային անոթային կապոցը, որը շարունակական է և ընդհանուր է տերևի և ցողունի համար:
Տերևի օնտոգենեզը վկայում է տերևի և ցողունի ընդհանուր բնույթի մասին՝ որպես մեկ ամբողջության՝ ընձյուղի մասեր։
Բողբոջի զարգացման պահից սկսվում է տերևի զարգացման արտաբողջական փուլը: Միեւնույն ժամանակ, տերեւների մակերեսը մեծանում է մի քանի հարյուր, նույնիսկ հազարավոր անգամ: Մակերեւութային աճը ձեռք է բերվում տերևի բջիջների մեծ մասը բաժանելով և դրանք երկարությամբ և լայնությամբ ձգելով:
Տերեւների կյանքի տեւողությունը կախված է գենետիկական եւ կլիմայական գործոններից։ Բարեխառն կլիմայի սաղարթավոր ծառերի և թփերի մոտ տերևների կյանքի արտաբողջային շրջանը 4-5 ամիս է։ Փշատերև բույսերի տերևները ապրում են երկուից վեց և ավելի տարի: Բայց ամեն դեպքում, տերեւների կյանքի տեւողությունը շատ ավելի քիչ է, քան բույսի առանցքային օրգանների կյանքի տեւողությունը։
Welwitschia mirabilis-ի հայրենիքը հարավ-արևմտյան Աֆրիկայի քարքարոտ անապատներն են: Այս թզուկ ծառն ունի երկար արմատ, հաստ և կարճ բուն (մինչև 50 սմ բարձրություն և մինչև 1 մ հաստություն) և երկու մեծ տերև, որոնք պահպանվում են ողջ կյանքի ընթացքում (մինչև 2000 տարի): Տերևների երկարությունը հասնում է 2-3 մետրի և անընդհատ աճում է հիմքում, վերևում մեռնում է։ Velvichia-ի համար խոնավության գրեթե միակ աղբյուրը խիտ մառախուղն է, որի խոնավությունը այս բույսը կլանում է տերևի երկու կողմերում գտնվող բազմաթիվ ստոմատների միջոցով (22200 ստոմատա 1 սմ 2-ի համար):
Տերեւների ակտիվ բաժանումը ճյուղերից՝ առանց նրա կենդանի հյուսվածքները վնասելու, կոչվում է ցրվածություն։ Ծառերի սեզոնային տերեւաթափը օրվա տեւողության փոփոխության արձագանքի արդյունք է։ Աշնանը տերևների անկումը զգալիորեն նվազեցնում է բույսի գոլորշիացման մակերեսը, որն անհրաժեշտ է աշնանը և ձմռանը ֆիզիոլոգիական խոնավության դեֆիցիտի պայմաններում։ Տերեւաթափի շնորհիվ կրճատվում է ձյան ծանրությունից ճյուղեր կոտրելու վտանգը։
Տերեւների մեծ մասում առանձնացնող շերտի առաջացումը տեղի է ունենում օնտոգենեզի ժամանակ։ Այս գոտում մեխանիկական հյուսվածքի քանակը կրճատվում է։ Թիլները ձևավորվում են շնչափողի տարրերում մինչև թափվելը, իսկ կալոզը նստում է մաղի տարրերում: Կտրման ժամանակ տեղի է ունենում բջջային թաղանթների ֆերմենտային ոչնչացում, ինչը հանգեցնում է բջիջների բաժանման միմյանցից: Մեմբրանների փոփոխությունները ներառում են միջնադարյան շերտի ցեմենտացման կարողության կորուստը (մասնակիորեն դրանից կալցիումի հեռացման պատճառով), բջջանյութի թաղանթների հիդրոլիզը և շնչափողի սկլերիֆիկացված տարրերի պատռումը:
Պաշտպանիչ շերտը կամ սպին առաջանում է թաղանթներում և միջբջջային տարածություններում պաշտպանիչ նյութերի, օրինակ՝ սուբերինի կամ վերքի մաստակի նստեցման արդյունքում։ ժամը փայտային բույսերպաշտպանիչ շերտը փոխարինվում է պերիդերմով, որը դրվում է պաշտպանիչ շերտի տակ և դառնում շարունակական ցողունի մնացած հատվածի հետ։
Տերեւների ցած թողնելը պարտադիր չէ, որ կապված լինի բջջային թաղանթների տարրալուծման գործընթացի հետ: Մեխանիկական անջատում առանց նախնական քիմիական փոփոխությունների կարելի է նկատել եղևնիից ասեղներ գցելու ժամանակ։
Տերեւների անկումից հետո տերեւի սպիները մնում են իրենց փնջի սպիներով։ Ճառագայթների սպիներ- սրանք անոթային կապոցների կոտրված ծայրերն են, որոնք տերևի հետքերից հասել են տերևի կոթուն մինչև այն ընկնելը:
տերևների հետքեր- սրանք ճյուղեր են ցողունի հաղորդիչ համակարգից մինչև տերևներ: Տերևի հետքը ձգվում է ցողունի կապոցի հետ իր միացման վայրից մինչև տերևի ելքը։ Մեկ տերևը կարող է ունենալ մեկ կամ մի քանի տերևի հետքեր:
Որովհետեւ ցողունը և տերևը ֆիլոգենետիկորեն ընդհանուր ծագում ունեն, տերևի հետքերի և ցողունի կապոցների միջև հիմնարար տարբերություն չկա: Համապատասխան տերմիններն ունեն նկարագրական, տեղագրական նշանակություն։
Հանգույցի շրջանում, որտեղ տերևի հետքը ցողունի կենտրոնական հաղորդիչ գլանից հեռանում է դեպի տերևի հիմքը, գլանում ձևավորվում է պարենշիմային գոտի՝ տերևի բացվածք կամ տերևի բացվածք։ Բույսից բույս տերեւային հետքերի եւ տերեւի բացերի թիվը տարբեր է:
Անգիոսպերմների առաջնային տիպը երեք կամ ավելի բացթողումներ ունեցող հանգույցն է: Անգիոսպերմի էվոլյուցիայի հիմնական ուղղությունը բնութագրվում է լակունների քանակի նվազմամբ։
Լակունները, միջուկը, միջֆասկուլյար գոտիները, հաղորդիչ համակարգը և հաղորդիչ համակարգի ծայրամասում տեղակայված պերցիկլը կազմում են բույսի առանցքային մասի (ցողուն և արմատ) ներքին միջուկը կամ. ստել.
Ստելների դասակարգումը հիմնված է զարգացման սկզբնական փուլում առանցքային օրգաններում հաղորդիչ և ոչ հաղորդիչ հյուսվածքների փոխադարձ դասավորության վրա։ Ստելի ամենապարզ տիպի մեջ, որը համարվում է առավել ֆիլոգենետիկորեն պարզունակ, հաղորդիչ հյուսվածքը կազմում է շարունակական սյունակ՝ քսիլեմը շրջապատող ֆլոեմով։ Սա նախաստել է։ Ցողունի նման կազմակերպումը կենսաբանորեն անկատար է, քանի որ հաղորդիչ և հիմքում ընկած հյուսվածքների միջև շփման մակերեսը փոքր է:
Ստելի հետագա էվոլյուցիան գնաց հաղորդիչ հյուսվածքների շփման մակերեսը հիմնականների հետ մեծացնելու ճանապարհով։ Դրան հաջողվել է երկու ճանապարհով. Մի դեպքում, ստելը խորը ելքեր է առաջացնում դեպի կեղևը և խաչաձեւ հատվածում աստղի տեսք ունի։ Սա ակտինոստել է։
Զարգացման երկրորդ՝ կենսաբանորեն ավելի խոստումնալից ճանապարհը պարենխիմային միջուկի հայտնվելն է ստելի կենտրոնում։ Սա սիֆոն է։
Տերեւների զարգացման հետ մեկտեղ առաջանում է հաղորդիչ և հիմնական հյուսվածքների ավելի սերտ կապ: Որքան շատ է տերևները, այնքան ավելի շատ է սթելը հագեցված հիմնական պարենխիմով: Ստելն ընդունում է ցանցի տեսք (հունարեն ցանց՝ dictuon)։ Սա դիկտիոստել է։
Ժամանակակից ծաղկող բույսերի ստելը էլ ավելի պարենխիմատացված է: Ստելը, որը հաղորդիչ թելերի և միջֆասկուլյար գոտիների համակարգ է, էուստելա է (հունարեն eu - լավ):
Միակոթուններում տերևների հետքերը հասնում են ցողունի կենտրոն, այնուհետև շեղվում են դեպի ծայրամաս։ Հետեւաբար, ցողունի խաչմերուկի վրա կապոցները կարծես ցրված են առանց որեւէ կարգի: Ատակտոս հունարենից. խառնաշփոթ. Այստեղից էլ առաջացել է միաշերտավորների ստելի անվանումը՝ ataktostele։ Նա բնութագրվում է բարձր աստիճան parenchymatization, երբ յուրաքանչյուր տերեւի հետք "
Արմատը առանցքային օրգան է, որն ունի ճառագայթային համաչափություն և երկարությամբ աճում է այնքան ժամանակ, քանի դեռ պահպանվում է գագաթային մերիստեմը։ Արմատը մորֆոլոգիապես տարբերվում է ցողունից նրանով, որ դրա վրա երբեք տերևներ չեն առաջանում, իսկ գագաթային մերիստեմը ծածկված է արմատային գլխարկով։ Արմատային սերունդ բույսերի մեջ պատահական բողբոջների ճյուղավորումն ու սկիզբը տեղի է ունենում էնդոգեն (ներքին)՝ պերիցիկլի (առաջնային կողային մերիստեմ) գործունեության արդյունքում։
Արմատային գործառույթներ.
1) Արմատը հողից ջուր է կլանում իր մեջ լուծված հանքանյութերով.
2) կատարում է խարիսխի դեր՝ ամրացնելով բույսը հողում.
3) ծառայում է որպես սննդանյութերի անոթ.
4) մասնակցում է որոշ օրգանական նյութերի առաջնային սինթեզին.
5) արմատային սերունդ բույսերում կատարում է վեգետատիվ վերարտադրության գործառույթ.
Արմատների դասակարգում. Ըստ ծագման, արմատները բաժանվում են հիմնական, պատահական և կողային: Սերմի բողբոջային արմատից առաջացող արմատը կոչվում է հիմնական; կոչվում են արմատները, որոնք առաջանում են բույսերի այլ օրգանների վրա (ցողուն, տերև, ծաղիկ): adnexal. Բուսական անգիոսպերմների կյանքում պատահական արմատների դերը հսկայական է, քանի որ չափահաս բույսերում (երկուսն էլ միաշաքիլեդոններ և շատ երկշիկավորներ) արմատային համակարգը հիմնականում (կամ միայն) բաղկացած է պատահական արմատներից: Ծիլերի հենակետային մասում պատահական արմատների առկայությունը հեշտացնում է բույսերի արհեստական բազմացումը՝ դրանք բաժանելով առանձին ընձյուղների կամ պատահական արմատներով ընձյուղների խմբերի։
Երիտասարդ ցողուններում պատահական արմատների սկզբնաղբյուրները ձևավորվում են էնդոգեն՝ միջֆասիկուլյար պարենխիմայի բջիջներից, հինում՝ առանցքային ճառագայթների կամբիումի մոտ գտնվող պարենխիմայից, այսինքն՝ պարենխիմալ ծագման կամբիումից։ Միաշերտ ցողունների պատահական արմատները առաջանում են կապոցները շրջապատող պարենխիմում:
Կողքարմատները ձևավորվում են հիմնական և պատահական արմատների վրա: Դրանց հետագա ճյուղավորման արդյունքում առաջանում են ավելի բարձր կարգերի կողային արմատներ։ Ամենից հաճախ ճյուղավորումը տեղի է ունենում մինչև չորրորդ կամ հինգերորդ կարգերը:
Մեկ բույսի բոլոր արմատների ամբողջությունը կոչվում է արմատային համակարգ.
ըստ ծագման:
հիմնական արմատային համակարգըզարգանում է բողբոջային արմատից և ներկայացված է հիմնական արմատով (առաջին կարգի) երկրորդ և հաջորդ կարգերի կողային արմատներով։ Միայն հիմնական արմատային համակարգը զարգանում է բազմաթիվ ծառերի և թփերի, ինչպես նաև միամյա և որոշ բազմամյա խոտաբույսերի երկկոտրուկների մոտ.
պատահական արմատային համակարգզարգանում է ցողունների, տերեւների, երբեմն՝ ծաղիկների վրա։ Արմատների այս ծագումը համարվում է ավելի պարզունակ, քանի որ այն բնորոշ է բարձրագույն սպորներին, որոնք ունեն միայն պատահական արմատների համակարգ։ Անգիոսպերմների մեջ պատահական արմատների համակարգը ձևավորվում է սերմերից, որում զարգանում է պրոտոկորմը (սաղմնային պալար), և հետագայում դրա վրա զարգանում են պատահական արմատներ.
խառը արմատային համակարգլայնորեն տարածված է ինչպես երկկոտորակների, այնպես էլ միաբնակարանների մեջ: Սերմերից աճեցված բույսում սկզբում զարգանում է հիմնական արմատի համակարգը, բայց դրա աճը երկար չի տևում. այն հաճախ դադարում է առաջին աճող սեզոնի աշնանը: Այս պահին հետևողականորեն զարգանում է պատահական արմատների համակարգ հիմնական ընձյուղի հիպոկոտիլային, էպիկոտիլային և հետագա մետամերների վրա, իսկ հետագայում՝ կողային ընձյուղների հիմքային մասում: Կախված բույսի տեսակից՝ դրանք սկսվում և զարգանում են մետամերների որոշակի հատվածներում (հանգույցներում, հանգույցների տակ և վերևում, միջհանգույցներում) կամ դրանց ողջ երկարությամբ։
Արմատային համակարգերի դասակարգում ձեւով.
Հիմնական արմատային համակարգը կոչվում է առանցքայինեթե հիմնական արմատը երկարությամբ և հաստությամբ նկատելիորեն գերազանցում է կողայինները.
Հիմնական և կողային արմատների նույն չափերով, արմատային համակարգով մանրաթելային. Խառը արմատային համակարգը նույնպես կարող է լինել առանցքայինեթե հիմնական արմատը շատ ավելի մեծ է, քան մյուսները, մանրաթելայինեթե բոլոր արմատները չափերով համեմատաբար հավասար են. Նույն տերմինները կիրառվում են նաև պատահական արմատների համակարգի համար։
Նույն արմատային համակարգում արմատները հաճախ կատարում են տարբեր գործառույթներ: Տարբերում են կմախքային արմատներ (կրող, ամուր, զարգացած մեխանիկական հյուսվածքներով), աճող (արագ աճող, բայց քիչ ճյուղավորվող), ծծող (բարակ, կարճատև, ինտենսիվ ճյուղավորվող)։



































- Չափը՝ 2 ՄԲ
- Սլայդների քանակը՝ 36
Ներկայացման նկարագրությունը ROOT AND ROOT SYSTEMS 1. Գործառույթներ և էվոլյուցիոն սլայդներ
ԱՐՄԱՏ ԵՎ ԱՐՄԱՏԱԿԱՆ ՀԱՄԱԿԱՐԳԵՐ 1. Արմատի ֆունկցիաները և էվոլյուցիոն ծագումը: 2. Արմատի առաջնային կառուցվածքը. 3. Երկրորդական արմատային փոփոխություններ. 4. Կողմնակի և պատահական արմատների ձևավորում: արմատային համակարգեր. 5. Արմատների մասնագիտացում և ձևափոխում.
Արմատը առանցքային օրգան է, որն ունի շառավղային համաչափություն և երկարությամբ աճում է անորոշ ժամանակով գագաթային մերիստեմի գործունեության շնորհիվ։ Տերեւները երբեք չեն հայտնվում արմատի վրա, իսկ գագաթային մերիստեմը միշտ ծածկված է գլխարկով։ Արմատի հիմնական գործառույթը ջրի և հանքանյութերի կլանումն է, այսինքն՝ բույսի հողի սնուցման ապահովումը։ Բացի վերը նշված հիմնական գործառույթից, արմատները կատարում են այլ գործառույթներ. նրանք ամրացնում են բույսը հողում, հնարավորություն են տալիս ուղղահայաց աճել և վեր թռչել; արմատներում տեղի է ունենում տարբեր նյութերի երկրորդական սինթեզ (ամինաթթուներ, ալկալոիդներ, ֆիտոհորմոններ և այլն); պահեստային նյութերը կարող են տեղակայվել արմատներում. արմատները փոխազդում են այլ բույսերի արմատների, հողի միկրոօրգանիզմների և սնկերի հետ:
Արմատները առաջացել են հողի մակերեսի վրա տարածված ռինոֆիտների տելոմներից։ Էվոլյուցիայի ընթացքում այս տելոմների որոշ ճյուղեր սկսեցին խորանալ հողի մեջ և արմատներ առաջացնել։
Արմատները հարմարեցված են հողի ավելի կատարյալ սնուցմանը։ Արմատների առաջացումը ուղեկցվել է դրանց ամբողջ կառուցվածքի խորը վերակառուցմամբ։ Նրանք ունեն մասնագիտացված հյուսվածքներ։ Հողից նյութեր կլանելու գործառույթը սկսեցին կատարել երիտասարդ արմատների վերջավորությունները։ Նրանք կենդանի բջիջներ են պահում մակերեսի վրա։ Այս բջիջները ձևավորեցին ֆունկցիոնալ առումով ամենակարևոր արմատային հյուսվածքը՝ ռիզոդերմը։
Հողից նյութեր կլանելու գործառույթը սկսեցին կատարել երիտասարդ արմատների վերջավորությունները։ Նրանք կենդանի բջիջներ են պահում մակերեսի վրա։ Այս բջիջները ձևավորեցին ֆունկցիոնալ առումով ամենակարևոր արմատային հյուսվածքը՝ ռիզոդերմը։ Այնուհետև, էվոլյուցիայի գործընթացում ներծծող արմատի մակերեսը մեծացավ երեք գործոնների պատճառով. 1) առատ ճյուղավորում և մեծ թվով ներծծող վերջավորությունների ձևավորում. 2) արմատների մշտական աճը և ներծծող վերջավորությունների տեղաշարժը դեպի հողի նոր տարածքներ. 3) արմատային մազերի ձևավորում.
Քանի որ արմատների աճը տեղի է ունենում խիտ հողում, դրա գագաթային մերիստեմը պետք է պաշտպանված լինի: Գագաթային մերիստեմը վնասվելուց պաշտպանված էր արմատային գլխարկի տեսքով: Արմատների տեսքը առաջացել է կլիմայի չորության աճով։ Ավելի չոր կլիմայի սկիզբը պատճառ է դարձել, որ ցամաքային բույսերը կցվեն ենթաշերտին և կլանեն ջուրն ու սննդանյութերը դրանից: Այնուամենայնիվ, էվոլյուցիայի ընթացքում տարբեր բույսերի տեսակների մեջ արմատի կառուցվածքը ավելի քիչ է փոխվել, քան ցողունը: Դա պայմանավորված է նրանով, որ հողային միջավայրում պայմաններն ավելի կայուն են, քան օդում։ Հետեւաբար, արմատը համարվում է ավելի «պահպանողական» օրգան, թեեւ այն հայտնվել է շատ ավելի ուշ, քան փախուստը: Արմատների ձևավորումը բույսերի կարևոր արոմորֆոզ է: Նրա շնորհիվ բույսերը կարողացան տիրապետել ավելի չոր հողերին և վեր բարձրացող մեծ ընձյուղներ առաջացնել։
Արմատի գլխարկի ամիլոպլաստների արձագանքը գրավիտացիայի: Շարժվող ստատոլիտներ խաղում կարևոր դերֆիտոհորմոնային գրադիենտներ ստեղծելու մեջ, որոնք ապահովում են արմատների ուղղահայաց աճը:
Ցորենի սածիլի արմատի կառուցվածքը (Triticum aestivum). Ա - արմատի կառուցվածքի դիագրամ; B - ռիզոդերմային և էկզոդերմային բջիջների տարբերակում: 1 - անցկացման գոտի, 2 - ներծծման գոտի, 3 - ձգվող գոտի, 4 - բաժանարար գոտի, 5 - արմատային մազ, 6 - արմատային գլխարկ:
Արմատի խաչմերուկ (a - միաշաքիլ, բ - երկշաքիլ բույս)
Առաջնային ծառի կեղևը առաջանում է պերիբլեմից: Նրա հիմնական զանգվածը կազմված է կենդանի պարենխիմալ բջիջներից՝ բարակ թաղանթներով։ Նրանց միջև ձևավորվում է միջբջջային տարածությունների համակարգ՝ երկարաձգված արմատի առանցքի երկայնքով։ Գազերը (CO 2) շրջանառվում են միջբջջային տարածություններով։ Գազերը անհրաժեշտ են կեղևի և ռիզոդերմիսի բջիջներում ինտենսիվ նյութափոխանակությունը պահպանելու համար: Կեղևի բջիջներում էներգետիկ նյութափոխանակությունը անհրաժեշտ է մի շարք կարևոր գործառույթներ կատարելու համար. 2) կեղևում սինթեզվում են տարբեր նյութեր, որոնք այնուհետև տեղափոխվում են այլ հյուսվածքներ. 3) կեղևի բջիջներում կուտակվում են պահուստային նյութեր. 4) կեղևում հաճախ հանդիպում են սնկերի հիֆեր, որոնք կազմում են միկորիզա։
Էնդոդերմի բջիջներն անցնում են զարգացման երեք փուլ. Կլանման գոտում էնդոդերմը գտնվում է առաջին փուլում։ Նրա բջիջների շառավղային պատերի մեջտեղում ձևավորվում են կասպարյան գոտիներ։ Կասպարյան գոտիները արգելափակում են նյութերի շարժումը բջջային թաղանթների միջով, այսինքն՝ ապոպլաստի երկայնքով: Երկրորդ փուլը կարելի է դիտարկել կողային արմատների գոտում։ Միաժամանակ բջջաթաղանթի ներքին կողմում հայտնվում է բարակ սուբերինային թիթեղ։ Այնուամենայնիվ, էնդոդերմը դեռևս ազատ է լուծույթներ փոխանցելու համար, քանի որ այն պարունակում է բարակ պատերով առանձին բջիջներ: Էնդոդերմի զարգացման երրորդ փուլը կարելի է դիտարկել միաշերտ արմատների գոտում։ Նրա բջիջների ներքին և շառավղային պատերը մեծապես հաստացել են։ Լայնակի հատվածների վրա այս բջիջները ունեն պայտաձև ձև: Անցակետեր չկան։ Հաստ պատերով էնդոդերմը պաշտպանում է հաղորդիչ հյուսվածքները և մեծացնում արմատների ամրությունը:
Կասպարյան գոտին անջրանցիկ պատնեշ է, որը ստիպում է ջուրը լքել ապոպլաստը և էնդոդերմի բջիջների թաղանթներով ներթափանցել սիմպլաստ:
Ջրի ներհոսքը հողից դեպի արմատ. ջուրը կարող է շարժվել ապոպլաստի և սիմպլաստի երկայնքով, մինչև այն հասնի էնդոդերմիս: Ապապլաստի երկայնքով հետագա շարժումն անհնար է:
Արմատի կենտրոնական գլանի կառուցվածքի տարբեր տեսակներ (առաջնային կառուցվածք)՝ Ա-դիարխիա, Բ-տրիարխիա, Գ-տետրարխիա, Գ-պոլիարխիա։ A-B տեսակներըբնորոշ է երկշաքիլավորներին, G - շատ միաշերտավորներում։ 1 - առաջնային ծառի կեղևի տարածք, 2 - առաջնային փեղկ, 3 - առաջնային քսիլեմ:
Հնարավոր է տարբերակել արմատի առաջնայինից երկրորդական կառուցվածքի անցման 4 փուլ. 2) ֆելոգենի առաջացում պերիցիկլով. 3) առաջնային ընդերքի թափում. 4) հաղորդիչ հյուսվածքների շառավղային դասավորության փոփոխությունը գրավով.
Անցում առաջնային արմատային կառուցվածքից երկրորդային, 11-ը` երկրորդական կեղև, 12-ը` ֆելոգեն, 13-ը` ֆելեմ:
Սիսեռի արմատում հաղորդիչ հյուսվածքների առաջնային տարբերակման սխեման 1 - քսիլեմի կենտրոնաձիգ տարբերակում, 2 - էպիդերմիս, 3 - առաջնային ծառի կեղև, 4 - էնդոդերմիս, 5 - առաջին տարբերակված քսիլոմային տարրեր, 6 - չտարբերակված քսիլոմային տարրեր, 7 - առաջին տարբերակված ֆլոեմ տարրեր. , 9 - գագաթային մերիստեմ, 10-արմատ գլխարկ:
Միակոթուների արմատի երկրորդական փոփոխությունները. Միաշերտ բույսերի ճնշող մեծամասնությունը պահպանում է առաջնային արմատային կառուցվածքը մինչև իրենց կյանքի վերջը։ Այնուամենայնիվ, արմատի շատ տարրեր ենթարկվում են lignification: Փայտային միաշերտավորների մեջ (արմավենիներ, dracaena, yucca) արմատային ծառի կեղևում առաջանում է մերիստեմային շերտ՝ պարենխիմային բջիջներից կամ պերցիկլից։ Դրանից կազմվում են փակ հաղորդիչ կապոցների շարքեր։ Անցկացնող կապոցների այս շարքից հետո առաջնային ծառի կեղևի պարենխիմայի ծայրամասային մասում առաջանում է կրթական հյուսվածքի նոր շերտ։ Այս մերիստեմային շերտը առաջացնում է անոթային կապոցների նոր շարք: Այսպիսով, արմատը խտանում է:
Հաճելի արմատները առաջանում են բույսերի տարբեր օրգանների վրա՝ ցողունների, տերևների և արմատների վրա։ Ցողունի վրա առաջացած պատահական արմատները կոչվում են ցողունային արմատներ, նրանք, որոնք առաջացել են արմատի վրա՝ արմատային արմատներ։ Կողային և պատահական արմատները էնդոգեն ծագում ունեն, այսինքն՝ դրված են ներքին հյուսվածքներում։
Արմատների կողային սկիզբը սկսվում է պերիցիկլի բջիջների բաժանմամբ: Այս դեպքում ստելի մակերեսի վրա ձևավորվում է մերիստեմատիկ տուբերկուլյոզ։ Մերիստեմատիկ տուբերկուլյոզի մի շարք բջիջների բաժանումներից հետո առաջանում է կողային արմատ։ Այն ունի իր գագաթային մերիստեմը և պատյանը: Կողային արմատի բողբոջը աճում է, ճեղքում է մայրական արմատի առաջնային կեղևը և շարժվում դեպի դուրս։ Սովորաբար կողային արմատները առաջանում են քսիլոմային տարրերի դեմ։ Հետեւաբար, դրանք դասավորված են արմատի երկայնքով կանոնավոր երկայնական շարքերով: Դրանք տեղի են ունենում կլանման գոտում կամ մի փոքր ավելի բարձր: Կողմնակի արմատների տեղադրումը տեղի է ունենում ակրոպետալ ձևով, այսինքն ՝ արմատի հիմքից մինչև դրա գագաթը:
Հարկավոր արմատները սովորաբար դրվում են մերիստեմատիկ գործունեության ընդունակ հյուսվածքներում՝ պերիցիկլում, կամբիումում, ֆելոգենում։ Կողային (և պատահական) արմատների էնդոգեն ձևավորումն ունի հարմարվողական արժեք։ Եթե ճյուղավորումը տեղի ունենար գագաթում, ապա արմատի առաջխաղացումը հողում դժվար կլիներ:
Դիխոտոմային ճյուղավորում մամուռի արմատային համակարգում (Lycopodium clavatum) 1 - ամենաբարակ արմատների իզոտոմային երկատված ճյուղավորում.
Միկորիզա՝ A - էկտոտրոֆիկ կաղնու միկորիզա, B, C՝ էնդոտրոֆիկ խոլորձի միկորիզա: Նոդուլներ լյուպի արմատների վրա