Корен и коренови системи. Образуването на странични корени
Филогенетично коренът стана по-късно стъблото и листа - във връзка с прехода на растенията към живота на земята и вероятно се случи от коренови подземни клонки. Коренът няма никакви листа или в определен ред на бъбреците. Той се характеризира с увеличаване на растежа по дължина, страничните клони възникват от вътрешните тъкани, точката на растеж е покрита с основен случай. Коренната система се образува през целия живот на растителния организъм. Понякога коренът може да служи като място на депозит в доставката на хранителни вещества. В този случай той е променен.
Видове корени
Главният корен се образува от зародишния корен по време на покълването на семената. Страничните корени отпътуват от него.
Корените под налягане се развиват на стъбла и листа.
Страничните корени са клони на корени.
Всеки корен (основен, страничен, очевиден) има способността да се разклонява, което значително увеличава повърхността на кореновата система и това допринася за по-добро укрепване на растението в почвата и подобряване на неговото хранене.
Видове коренови системи
Има два основни вида коренови системи: пръчка, с добре развит главен корен и урина. Коренната система на урината се състои от голям брой очевидни корени, същата най-голяма. Цялата маса на корените се състои от странични или очевидни корени и има външен вид на лоб.
Силната разклонена коренова система образува огромна абсорбираща повърхност. Например,
- общата дължина на корените на зимния ръм достига 600 км;
- дължината на коренните косми е 10 000 км;
- общата повърхност на корените е 200 m 2.
Това е много пъти повече от площта на горната маса.

Ако растението е добре изразено от главния корен и се развиват очевидните корени, се образува кореновата система от смесен тип (зеле, домати).
Външната структура на корена. Вътрешната структура на корена
Root zones.

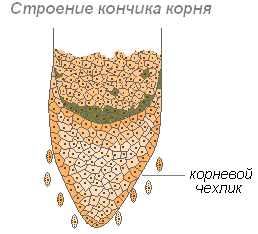
Корен калъф
Коренът расте с дължина с върха, където се намират млади клетки на образователната тъкан. Нарастващата част е покрита с основен случай, който защитава върха на корена от увреждане и улеснява напредъка на корена в почвата по време на растежа. Последната функция се извършва поради свойството на външните стени на коренния случай, за да бъде покрит със слуз, което намалява триенето между корена и почвените частици. Може дори да избута частиците на почвата. Клетките на основния случай са живи, често съдържат грама нишесте. Целзите постоянно се актуализират поради разделение. Участва в положителни геотропни реакции (посока на растежа на корен към центъра на Земята).
Клетките на зоната на разделяне са активно разделени, дължината на тази зона в различни видове и различните корени от същото растение не е същото.
Зоната на разделението се намира зад зоната на разделяне (зона за растеж). Дължината на тази зона не надвишава няколко милиметра.
Тъй като линейният растеж е завършен, се образува третият етап от образуването на корените - се образува диференциация, зона на диференциация и специализация на клетки (или зона на коренни косми и засмукване). В тази зона външният слой на епибломи (ризодетерм) с коренни косми, слой от първична кора и централен цилиндър са вече разграничени.
Структурата на коренните косми
Коренните косми са силно удължена лекота на външни клетки, покриващи корена. Броят на коренните косми е много голям (1 mm 2 от 200 до 300 косми). Дължината им достига 10 мм. Косата се образуват много бързо (в млади ябълкови разсад за 30-40 часа). Коренните косми са краткотрайни. Те умират след 10-20 дни, а на младата част на корена растат нови. Това осигурява развитието на корена на нови почвени хоризонти. Коренът непрекъснато нараства, образувайки нови и нови секции от коренови косми. Коса могат не само да абсорбират готовите разтвори на вещества, но и да допринесат за разтварянето на определени вещества на почвата, и след това да ги смучат. Коренната секция, където коренните косми изчезнаха, известно време е способен да всмуква вода, но след това да бъде покрит с щепсел и загуби тази способност.
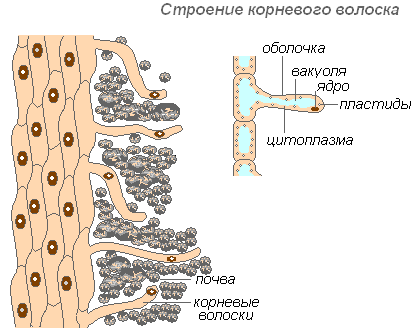
Обвивката на космите е много тънка, което улеснява усвояването на хранителните вещества. Почти цялата клетка за коса заема вакуория, заобиколен от тънък слой цитоплазма. Ядрото е в горната част на килията. Около клетката се образува случай на лигавицата, който насърчава лепене на коренни косми с почвени частици, което подобрява контакта им и увеличава хидрофилността на системата. Абсорбцията допринася за разделянето на киселини (въглища, ябълка, лимон), които разтварят минерални соли.
Коренни косми Играйте и механична роля - те подкрепят върха на корена, който преминава между почвените частици.
Под микроскопа на напречното сечение на корена в смукателната зона се вижда своята структура в клетъчните и тъкани. На повърхността на корена - Risoderma, под него - кора. Външният слой на кората е екзодерма, навътре от нея - основната паренхима. Неговите тънкостенни живи клетки изпълняват пенлива функция, хранителните вещества в радиалната посока се извършват - от всмукване на дървесни съдове. В тях се случва синтез на редица жизнени органични вещества. Вътрешният слой на кора е ендодерма. Разтворите на хранителни вещества, идващи от кора в централния цилиндър през клетките на ендодермата преминават само през протопластичните клетки.

Кората заобикаля централния цилиндър на корена. Той граничи с слой от клетки за дълго време, запазвайки способността да се разделят. Това е перикола. Периколесните клетки започват странични корени, очевидни бъбречни и средни образователни тъкани. Вътрешно от перикола, в центъра на корена, има проводими тъкани: LOB и Wood. Заедно те образуват радиален проводим лъч.
Проводимата коренова система провежда вода и минерални вещества от корена в стеблото (възходящ ток) и органични вещества от стъблото към корена (надолу). Състои се от съдови влакнести лъчи. Основните компоненти на лъча са наводненията на етажа (веществата се преместват в корена) и ксилари (за които вещества се движат от корена). Основните проводими елементи на етажа са тръби за сито, ксилеми - трахея (съдове) и трахедис.
Процеси на корен живот
Воден транспорт в корена
Абсорбцията на вода с коренни косми, изработени от почвен хранителен разтвор и я води в радиалната посока през клетките на първичната корка през пропуснатата клетки в ендодерм към ксилола на радиалния проводящ лъч. Интензивността на абсорбцията на вода коренни косми се нарича смучене сила (и), е равна на разликата между осмотичното (p) и турне (t) налягане: s \u003d p-t.
Когато осмотичното налягане е равно на турнето (р \u003d t), след това s \u003d 0, водата престава да тече в коренната козина. Ако концентрацията на почвените хранителни вещества е по-висока, отколкото в клетката, водата ще излезе от клетките и ще дойде плазмолиза - растенията са покрити. Такъв феномен се наблюдава при условия на сухота на почвата, както и с неограниченото производство на минерални торове. Вътре в кореновите клетки, причиняването на якостта на корена се увеличава от ризодермата към централния цилиндър, така че водата се движи по градиента на концентрацията (т.е. от мястото с по-висока концентрация на място с по-малка концентрация) и създава коренов натиск който повдига водния стълб от съдовете на ксилемите чрез образуване на възходящ ток. Това може да бъде открито на пролетни чупливи куфари, когато събират "сок" или върху нарязани пънове. Разширяване на вода от дърво, пресни пънове, листа, наречени "плач" растения. Когато листата цъфтят, те също така създават смукателна сила и да привличат вода - във всеки съд се образува непрекъсната водна колона - капилярно напрежение. Кореновият натиск е долния двигател на водния ток, а смущаващата сила на листата е горната част. Това може да бъде потвърдено с помощта на прости експерименти.
Абсорбция на вода Корени
Предназначение: Разберете основната функция на корена.
Какво да правя: Растението, отглеждано на мокри дървени стърготини, ще отмени с кореновата си система и ще го сложи в чаша с вода с корените си. На върха на водата, за да го предпази от изпаряване, тънък слой растително масло и споменава нивото.

Какво се наблюдава: В ден или две водата в резервоара намалява под марката.

Резултат: Следователно корените на водата и го подадоха до листата.
Все още можете да направите един опит, доказващ хранителен корен.
Какво да правя: Продаден в завода, стъблото оставяйки височина на пяната от 2-3 см. На порети носят гумена тръба с дължина 3 cm, а на горния край постави на извита стъклена тръба с височина 20-25 cm.
Какво се наблюдава: Водата в стъклена тръба се издига и излиза.
Резултат: Това доказва, че водата от корена на почвата е гадно в стеблото.

Има ли температура на водата върху интензивността на засмукване от корена на водата?
Предназначение: Разберете как температурата засяга работата на корен.
Какво да правя: Една стъкло трябва да бъде с топла вода (+ 17-18ºС), а другата със студ (+ 1-2ºС).
Какво се наблюдава: В първия случай водата се откроява изобилно във втория - има малко или напълно спряно.
Резултат: Това е доказателство, че температурата силно засяга работата на корен.

Топлата вода се абсорбира активно от корените. Кореновият натиск се повишава.

Студената вода се абсорбира слабо от корените. В този случай корекционният натиск намалява.
Минерално хранене
Физиологичната роля на минералите е много голяма. Те са основата за синтеза на органични съединения, както и фактори, които променят физическото състояние на колоидите, т.е. пряко засягат метаболизма и структурата на протопласта; изпълнява функцията на катализатори на биохимични реакции; засягат клетките на клетките и пропускливостта на протоплазма; са центрове на електрически и радиоактивни явления в растителните организми.
Установено е, че нормалното развитие на растенията е възможно само в присъствието на три неметали в хранителния разтвор - азот, фосфор и сяра и - и четири метали - калий, магнезий, калций и желязо. Всеки от тези елементи има индивидуална стойност и не може да бъде заменена от друга. Това са макроелементи, концентрацията им в завода е 10 -2 -10%. За нормалното развитие на растенията са необходими микроелементи, концентрацията на която в клетката е 10 -5 -10 -3%. Това е борен, кобалт, мед, цинк, манган, молибден д-р. Всички тези елементи са в почвата, но понякога недостатъчни. Следователно минералните и органичните торове допринасят за почвата.
Заводът обикновено расте и развива в случай, че в околната корена ще съдържа всички необходими хранителни вещества. Такава среда за повечето растения е почвата.
Дишащи корени
За нормален растеж и развитие на растението е необходимо да се стигне до корена свеж въздух. Проверка, нали?
Предназначение: Дали въздухът се нуждае от корена?
Какво да правя: Вземете два идентични съда с вода. Във всеки кораб ще поставим развитието на разсад. Водата в един от съдовете всеки ден е наситен с въздух с пулверизатор. На повърхността на водата във втория съд тънък слой от растително масло, тъй като той забавя въздушния поток във водата.

Какво се наблюдава: След известно време растението във втория кораб ще престане да расте, ще обитава и в крайна сметка ще умре.

Резултат: Смъртта на растението се дължи на липсата на въздух, необходима за дишането на корена.
Модификации на канала
Някои растения имат резервни хранителни вещества в корените. Те натрупват въглехидрати, минерални соли, витамини и други вещества. Такива корени растат силно с дебелина и придобиват необичаен външен вид. Коренът и стремежа участват в образуването на корен.
Корени
Ако в главния корен се натрупват резервни скокове и в основата на главното бягство стебло, са оформени корените (моркови). Растения, образуващи корените, предимно здрач. През първата година от живота те не цъфтят и се натрупват в вкоренени много хранителни вещества в root. Във втория - те бързо цъфтят, използвайки натрупаните хранителни вещества и образуват плодове и семена.
Коренови клубени

Georgine, резервни вещества се натрупват в очевидните корени, образувайки коренови клубени.
Бактериални клубени
Страничните корени на детелината, лупина, люцерна се промениха особено. Бактериите се установяват в млади странични корени, което допринася за абсорбцията на газообразен азот на почвен въздух. Такива корени придобиват външния вид на клубена. Благодарение на тези бактерии тези растения са способни да живеят върху бедни азотни почви и да ги направят повече плодородни.
Пешеходци

Растената, която растат в приливната и приливната зона, развива корените на развалина. Те държат високо над водата в хъскивите орливни земни големи плодотворни издънки.
Въздух

В тропическите растения, живеещи върху клоните на дърветата, се развиват въздушни корени. Те често се срещат в орхидеи, Бромелиев, в някои пачове. Въздушните корени висят свободно във въздуха, без да достигат земята и да поглъщат влагата върху тях от дъжд или роса.
Прибиране

На луковични и туберюкуковични растения, например, кръстосани, сред многобройни филиментални корени, има малко по-дебел, така наречен ревляк, корени. Намаляване, такива корени нарисуват Tererukovitsa по-дълбоко в почвата.
Стълбове

Ficus развива стълбове, или корени.
Почвата като коренови местообитания
Почвата за растенията е средата, от която получава вода и батерии. Количеството минерални вещества в почвата зависи от специфичните характеристики на майчината скала, дейностите на организмите, от жизнената дейност на самите растения, върху вида на почвата.
Частиците на почвата се конкурират с корени за влага, задържайки го с повърхността си. Това е така наречената свързана вода, която е разделена на хигроскопична и филма. Тя се държи от силите на молекулярно привличане. Наличната влагодарна инсталация е представена с капилярна вода, която е концентрирана в малки пори на почвата.
Между влага и въздушна фаза, антагонистичните отношения се развиват. Колкото по-голям е в почвата на големи, толкова по-добър газ режим на тези почви, толкова по-малко влага държи почвата. Най-благоприятният водоустойчив режим се поддържа в структурни почви, където водата и въздухът се намират едновременно и не се намесват помежду си - водата запълва капилярите в структурните звена и въздухът е големи пори между тях.
Естеството на взаимодействието на растението и почвата е до голяма степен свързано с абсорбционния капацитет на почвата - способността за задържане или свързване на химични съединения.
Почвената микрофлора разлага органични вещества по-прости съединения, участва в образуването на структурата на почвата. Естеството на тези процеси зависи от вида на почвата, химичен състав Растителни остатъци, физиологични свойства на микроорганизми и други фактори. При образуването на структурата на почвата, почвените животни участват: звънене червеи, ларви на насекоми и др.
В резултат на съвкупността от биологични и химични процеси в почвата се образува сложен комплекс от органични вещества, който се комбинира с термина "хумус".
Метод на водна култура
В кои соли има растение и какво влияние те имат върху растежа и развитието на него, той е създаден по опит с водните култури. Методът на водните култури е отглеждането на растения, а не в почвата, но във воден разтвор на минерални соли. В зависимост от целта на целта в експеримента е възможно да се изключи отделна сол от разтвора, да намали или увеличава неговото съдържание. Установено е, че торовете, съдържащи азот, допринасят за растежа на растенията, съдържащи фосфор - ранното зреене на плодове и съдържащ калий - най-бързият изтичане на органични вещества от листата към корените. Във връзка с това, азотът на тора се препоръчва да бъде направен преди засяването или през първата половина на лятото, съдържаща фосфор и калий - през втората половина на лятото.
Използвайки метода на културата на водата, е възможно да се установи не само нуждата от растения в макроелементи, но и да се разбере ролята на различни микроелементи.
Понастоящем има случаи, когато растенията се отглеждат по методи на хидропоника и въздухоплавателни средства.
Хидропоника - растящи растения в съдове, пълни с чакъл. Хранително вещество необходимите елементи, Той се сервира в съдовете по-долу.
Самолетът е въздушната култура на растенията. В този метод кореновата система е във въздуха и автоматично (няколко пъти в рамките на час) се напръсква със слаб разтвор на хранителни соли.
Организация
Организация - Описание на органите на живия организъм.
Орган - част от тялото, диференцирана морфологично и специализирана функционално. В съответствие с двете основни функции: поддържане на живота на това растение и го възпроизвежда в редица последващи поколения - разпределят вегетативни и генеративни (репродуктивни) тела.
Първо, ние ще изучаваме вегетативните органи, образуващи системата за бягство и кореновата система. Коренната система е коренът.
Корен
Коренът на еволюцията се появява по-късно избяга и идва от ризоидите на първите земи - ринофит.
Различните части на кората изпълняват неравномерни функции и се характеризират с определени морфологични характеристики. Тези части са получили името на зоните. Зоните на младия корен: зоната на разделяне, зоната на растеж или разтягане, зоната на абсорбция или всмукване и площта на стопанството.
Извън апикалната меристем е покрита с основен случай. Жива паренхимна клетка произвеждат слуз. Лигавицата е силно хидратиращ полизахарид на пектинската природа. Тя се натрупва в мехурчета на докяза, която се слива с плазмолмата, да го освободи в пространството между плазмолема и клетъчната обвивка. Тогава слузът отива на повърхността на клетъчната обвивка, където образува малки капки.
Благодарение на това лигавицата, почвените частици се придържат към корена и коренните косми. Слузът служи като абсорбираща повърхност, която изключва йони. Лигавицата покритие от корена вредни вещества Почвата и сушенето създава благоприятни условия за уреждане на полезни бактерии.
Като корен и бутане на основния случай напред, клетките на периферията на кавалерията са изброени. Продължителността на живота на клетките от появата на обяда е 4-9 дни, в зависимост от дължината на кавалерията и вида на растението.
Коренният случай контролира георезба на корена. Мястото на възприемане на тежестта в случая - централната колона на клетките - Колумела, при която амилопластите действат като стафити (гравитационни рецептори).
APICAL MERISTEM се състои от малки (10-20 микрона в диаметър) на многостранни клетки с гъста цитоплазма и големи ядра.
Споделете растения монониален тип корен appex.
В корените на растенията за семена са описани два основни вида апикални организации. В един от тях всички области имат общи инициали. Това е отворен тип апикална меристем, която вероятно е филогенетично примитивна. Във втория тип, всяка от трите части, т.е. Централен (или проводим) цилиндър, първичен кората и корен случай, имат своя собствена първоначална. Това е затворен тип апикал.
Игноричност, отговорна за организирането на клетъчния растеж на първите етапи, главно губят митотична активност при следващите етапи на растежа на корените. Вместо това, митотичната активност показва клетки, разположени донякъде по-дълбоко в кореновото тяло. Тези наблюдения служат като причина за формирането на теорията на Центъра за почивка в апикалната мериста. Според тази теория най-дисталните клетки (дистални - най-отдалечените от централната част на органа: долния край на корена) в кореновото тяло (първия първоначален пелс и перибъл) рядко се споделят, имат за същото Размери и ниски скорости на синтез на NK и протеин. Центърът за почивка има полусферична или дискова форма, варира в количеството в зависимост от размера на корена. Клетките на центъра за почивка са разделени около 10 пъти по-бавни от съседните клетки.
В корените, евентуално повредени, центърът за почивка е в състояние да възстанови мири. Тези., Центърът за почивка играе значителна роля при формирането на структурата и в развитието на корена.
Характеристика на върха на корена е поставянето на надлъжни редове клетки, произхождащи от апикалната мериста.
Слоеве на APICAL MERISTEM ROOT:
Дерматоген. (Гръцка дерма - кожа, раждане на геноси) - външен слой на топ месистемата, е диференциран в Risoderma. Дерматогенни клетки на правилната форма.
Плерома (Гръцки. Плерома е пълнене) - централната част на елита meristem се диференцира в клетките на централния цилиндър. Клетките се удължат по оста.
Лед (Гръцки пепел - покритие) - слой от топ месистка между дерматоген и плола, диференцирани в клетките на първичната кора.
Първичната корка е диференцирана на нивото на разтягащата зона. В зоната на урината основната кора е отчетена за голяма част от напречното сечение. Той се нарича "първичен" поради произхода от първичната меристема, "кората" - поради периферно местоположение.
Външните клетки на първичната кора, разположени под ризодерма, се наричат \u200b\u200bанодерма. Вътрешният слой на първичната кора - ендодерма.
По-голямата част от основната кора е мезодерм. Мезодермата се образува от паренхимни клетки, разположени в насипно състояние, което допринася за аерацията на кореновите клетки.
Корените на гласуваното и най-дикотислодно рано нулиране на основната кора. Те имат първични клетки на кората, остават паренхим. За разлика от това, основната кора на една спалня остава през целия живот на корена, а клетките му образуват вторични лигнирани черупки.
Центърът на корена заема аксиален цилиндър - Stele. Започва да се разграничава в зоната на растеж. Образуването на аксиалния цилиндър започва с образуването на външния му слой - перикола. Перикола се отнася до първичните странични меристеми, клетките отдавна запазват meristematic активност. Обикновено перичкуването е представено с един клетъчен слой, въпреки че се случва и многослойно.
За повечето семена растения в перикола се полагат странични корени, възникват подход на очевидни бъбреци. При вида със вторичен растеж, периколката участва в образуването на Камбия и дава началото на първия слой на гълъба.
Под перикола се полагат клетки на прондмбия, които водят до първичен етаж, по-късно - първичен xylem. Броят на лъчните ксиславе варира от различни видове от две, три, четири до няколко. Съответно, броят на лъчите на хълмните корени се нарича диарх, трийчки, тетрархи, полиархи. Тетрархите корени се считат за първични. При монокони - Полярна стел.
По този начин, общ план Първичната структура на корените и морфологичната диференциация на тъканите показват висок стадий на специализация и приспособление на корена към функциите на засмукване и провеждане на вода и минерални вещества.
Образуването на странични корени
Страничните корени се появяват върху периферията на централния цилиндър на различни разстояния от апикалната меристи. Като Те са положени в дълбоко разположени коренови тъкани, техният произход се нарича ендогенен (с гръцки. "Роден вътре"). Страничните корени на гласуваните и покрити мостове са положени в перикола. Периколесни клетки, участващи в образуването на странични корени - кореновата дъга.
Няколко слоя кореноплодни клетки могат да образуват и да endoderma.
В корена на диарета, страничният корен е положен между плаващия и Ksiloi, в tryearn, тетраррхеном - противоположния Xleema, в монокоания полиард - срещу платието. Периколесните клетки са разделени различни области. В резултат на това се образува малък корен туберкул. Чрез увеличаване по размер, коренът на корена преси на ендодермата. Но мембраните на ендодерм са твърди. Ето защо е подчертана ендодермата. Измъчването на Endoderma се нарича корен джоб.
Приоритетът на корена преминава през първичната кора, закрепвайки ензимите, които унищожават мозъчните клетки по пътя им. В ранните етапи на развитие този пол образува основния случай, апикални и първични меристи. С течение на времето, поради диференциацията на паренхимните клетки в елементите на ксиславе и флора, централните цилиндри на страничния и родителския корен са свързани заедно.
Монокоторната първична коренна структура се поддържа през целия живот и вторичните тъкани не възникват. С възрастта, размерът на клетките може да се увеличи, клетъчните мембрани са удебелени, но общото "план" на коренната структура остава непроменено.
Вторичните промени се извършват при гласуване и дикотяница, а вторичната структура на корена се образува. Образуването на вторичната структура на корена е свързано предимно с дейностите на Камбия. Cambier се формира чрез разделяне на тези сканиращи клетки, които остават недиференцирани между първичния етаж и първичния ксилен. След това периколите, локализирани извън издатините на KSIL, също показват камбиална активност.
Камбий отлага вторична ксилемна клетка към центъра, до периферията - клетките на вторичния етаж. Така се образуват проводящи греди. Те се образуват толкова, колкото и лъчите на първичния ксиле. Под налягането на нарастващите вторични тъкани основните килии на етаж са сплескани. Първичният Xylem запазва централното положение и се вижда като звездичка. С течение на времето и първият Xylem също е извършен.
С вторичната структура на корена, първичните тъкани не функционират като проводими елементи. Токът на веществата в двете посоки се извършва според вторичните тъкани.
Cambier, който произхожда от перикола, образува широки паренхимни лъчи, които се поставят между отворени обезпечения лъчи. Това са първични сърдечни лъчи. Те осигуряват физиологичното свързване на централната част на корена с първична кора. По-късно, по-тесните вторични лъчи могат да бъдат положени заедно, свързващи вторичния ксисла и флорем.
Вторичните промени се срещат не само в централния цилиндър. Поради повишаването на дебелината на тъканите на тъканите, първичната кора се премества навън, избухва и се нулира заедно с ендодермата.
Във външната част на перикола се връща началото на гълъба и излага външната страна на Fellem, и навътре - многослойна гора. Така, корените на две дебелото черво, които имат вторична структура, покрита с перидерма.
В многогодишните корени на гълъба възникват в дълбините на корена. След това коренът, като стеблото, формира ритид.
Важен екологичен аспект на рециклиран корен Това е способността на корените на различни растения от един и същ вид към естествения пожар. Когато корените влизат в контакт с друг, те се комбинират благодарение на вторичния растеж. Когато корените са бързо, се създава непрекъснатост на проводимата система между двете пресичащи се партньори. Благодарение на стрелбата на корените, много дървета са взаимосвързани и ако някои от тях са изсечени, тогава пъпките остават живи за дълго време. Фантастиката на корените допринася за бързото предаване на инфекция от едно дърво на друго.
Диференциация на корените в кореновите системи В различни форми. Най-важна е диференциацията от върха, страничните и очевидните корени.
Корените, възникнали в апикалните меристи с по-високи спорове, се отличават в специална група. Предполага се, че те са най-архаичната група от корени и запазват най-голямото сходство с основните корени, които са част от една клонова система от режийни и подземни оси. Корените, включени в тази група, се наричат \u200b\u200bаплициен.
Въз основа на последователността на кореновите системи в процеса на еволюцията и принадлежащи към големи системни групи, можете да планирате три еволюционно ниво на кореновите системи.
I. Корените системи с по-високи спорове са представени изключително от аплициейни корени системи, които последователно се заменят като основен механизъм за актуализиране в кореновите системи. Разклоняване на върха или отстрани и е възможно и двата вида разклонения да възникнат независимо един от друг. Няма метаморфози и корени с тясна специализация.
P. Системата за вкореняване на подаръците се отличава с присъствието на главния корен, който може да бъде свързан с появата на семена и биполярно ембрион с зародителен корен. В механизма за актуализиране са включени не само страничните корени на подчинените поръчки, но и ризоко-коогенични уреди (т.е. очевидните корени на корените, очевидните корени не са получени от гласуваното забележително развитие). Специализирани и метаморфни корени са единични.
Sh. Коренната система на покрита, подредена се характеризира със силното развитие на очевидните корени, както и метаморфози и корените с тясна специализация. Механизмите за обновяване са разнообразни и ефективни.
Метаморфоза на кореновите системи
Метаморфозата на кореновите системи е комбинация от анатомични и морфологични характеристики, разработени от филогенетично и наследствено фиксирани и причинени от промяна във физиологичните функции. Метаморфоза (метаморфоза, гръцко-трансформация) - резултатът от промените, които се появяват на генетичното ниво и които водят до морфологични и функционални пренареждания на органа, системата на органите или тялото като цяло.
I. Метаморфоза, свързана с изпълнението на нетипични функции:
1) дишащи корени;
2) въздушни корени на епифите;
3) скитащи корени;
4) Корените на архива.
P. Метаморфоза, свързана с хиперфункция:
1) корен;
2) коренни конуси;
3) коренови възли;
4) Микориза.
Корените са характерни за двугодишни двустранни растения. Това е условно име. Те се развиват от вегетативни тела и нямат отношение към плодовете. Дъното на стъблото и коренът участват в образуването на корена.
През първата година от живота растителната растителност се образува коренът. За втората година се образуват цветя, плодове, семена. Това означава, че коренът на веществата е органът на резервите, който позволява на растението да прехвърля период на почивка и да завърши развитието на следващата година.
Видове rooteplood:
1.Монкамбил.
Източникът е коренът с аксиален цилиндър. Образуването на корена на корена започва с вторичните промени, свързани с присвояването и работата на един камбиален пръстен и по-нататъшна панеримализация.
Ние знаем, че Касбий вътре отложенията вторият ксислам, външен - вторичен флолам. Анатомичните различия се отнасят до мястото на развитие на математа за съхранение. Ако силното развитие на паренхима се появява на сношението, се образува PhLoem вид корен - вид моркови. Ако XILEM е Xylem тип - тип ряпа.
2. Полимбиален тип rooteplood.
Положен е един типичен камбиален пръстен, останалите са допълнителен перицикличен произход. Те могат да бъдат до 25. Тези допълнителни пръстени на камбия образуват отворени обезпечения проводими греди. По-нататъшният пръстен от първия камбанен пръстен, толкова по-голям произвежда pourens, той произвежда. Пример за поликомбиален корен корен - цвекло.
В корени моно и поликабиални корени, първичната кора е дълбоко умира и лишове. Покривната кърпа е periderma.
По този начин общият знак на всички корени е наличието на голям брой искрящи паренхим, проникнати с проводими тъкани. Тясната връзка между проводимите тъкани и запитването се постига чрез различни модификации на вторичния растеж.
В някои растения корени от отглеждане се формират от странични и очевидни корени под формата на коренови конуси. Коренните конуси на една спалня (Lubbar) Busty, имат първична структура; Разворният - камбанален, има вторична структура. Пример: Чипове от корен от тип Dahliam.
Коренните конуси по цялата повърхност образуват очевидните корени, а в горната част - очевидните бъбреци и следователно са органи на вегетативно размножаване.
Известно е, че растежът на растенията най-често ограничава недостатъчното осигуряване на фиксирания им азот. Азот под формата на стабилна молекула N2 е 80% от атмосферата. Преди фиксиране (с възстановяването на N2 до амоняк NH3), тази молекула трябва да се раздели. Фиксирането на азот се извършва от някои свободни бактерии, които консумират органичното вещество на почвата. Някои бактерии се заселват на корените на висшите растения. Растението "привлича" бактерии, използвайки органични коренови секрети. И микроорганизмите на свой ред осигуряват фиксирано азотно растение. Такава взаимно намалена асоциация на два организма се нарича симбиоза. Най-често корените образуват асоциации с бактерии за фиксиране на азот на рода ризобий. Повече от 60 семейства хранителни и моноциклични растения са в състояние да влязат в симбиоза с бактерии. Всеки тип растение има свой собствен симбиотичен ризобий. Взаимното разпознаване на растения и бактерии се извършва чрез закрепване на специален протеин - лектин, разположен на повърхността на трихобластите, до определена бактерия. Коренни косми под влиянието на хормони на растежа, разпределени от бактериалната деформация. Бактериите проникват в тези извити клетки. Вътре в клетките на бактерията многократно, образувайки инфекциозна нишка, сключена в случай от венците. Тази нишка прониква дълбоко в корена. Резултатът от такова проникване на бактериите е силният растеж на клетките на клетъчните мезодерм, което води до образуването на клубена. Самата външна част на сламата остава много време, а бактериите се поставят във вътрешната част на клубена. Самата бактериална тъкан е представена от малки клетки с вискозна цитоплазма и огромен брой бактерии. Бактериалната тъкан е заобиколена от проводящи лъчи, които са свързани с централния цилиндър на корена. Тези. Има транспортна система за обмен на хранителни вещества между бактерии и растения. Азотните бактерии имат благоприятен ефект върху храненето с азот на растенията и върху плодородието на почвата.
Физиологичната активност на корена, свързана с осигуряването на растения с вода и минерали, може да бъде засилен от своята симбиотична асоциация със специфична MUSA. Такава тясна взаимосвързана симбиоза на корените и гъбите и е микориза. Mikoriza е известен с повечето съдови растения. Само няколко цъфтящи семейства не го образуват или образуват рядко, например, зеле, умира.
Mycorris гъби играят ключова роля в минералното хранене на съдовите растения. Значителна роля на Микорижа в усвояването и прехвърлянето на фосфор.
С двустранен транспорт (движещи се фосфати от гъби до растения и въглехидрати в обратна посока) загуба и абсорбция се появяват в апопластичната зона между плазмените мембрани на растенията и гъбичките.
Mycorrisian гъби използват тези източници p, които обикновено не са достъпни за растенията. Микоридж също допринася за усвояването на ZN, MN, CU растения. Тези елементи в почвата са слабо втечнени и следователно около коренните косми се образуват бързо от зоните, изчерпани от тях. Мрежата от химикали на микорисовите гъби се разпространява до няколко сантиметра от всеки корен и по този начин увеличава повърхността на абсорбцията на корените.
Разграничават се следните видове микорнис:
1) Превъплъстяване: гъбата покрива целия връх на корена с дебел случай и прониква в междуклазерите;
2) Ендомикоридност: гъбата е вградена вътре в клетките.
Екстраторизмът е характерен за някои семейства на дървета и храсти (бор, ява, бук). Екстрамикорозата се образува на млади, все още не тествани корени. Акстрасорните корени са къси, разклонени и вид на поглъщане.
Тези корени, развитието на кореновите косми се потискат и обемът на апикалния меристи и корен може да бъде намален. Функцията на коренните косми се извършва от GIF файлове.
Ендомикоризираните корени във формата са сходни с обикновените корени, но се различават от тях по-тъмно в тъмното. Гъби Gifs проникват в мозъла на коренната корка, където се образуват весикули (подуване) и фъфюслас (клони). Следователно, ендомикорозата често се нарича везикуларна въоръжа. При определени условия, GIF файловете на гъбата могат да "усвоят" от клетки. Коренните косми на корените се запазват. Това е най-често срещаният тип микориза. Той се среща приблизително 80% от всички съдови растения, а именно в билкови растения, включително зърно; При храсти, дървета (ябълкови дървета, кленове ...). В младото тополово растение, например, се образува везикуларна арматура-ендомикороза и като е съгласна - покриваща ектомикороза.
За Heers, Ericaceae се характеризира с Ерицидо-Арбутоид Микориза. Гъбата около корена образува мощно натрупване на GIF и тънките странични Gifs проникват в сандъка на корекса. Тези минерални гъби значително намаляват токсичността на почвата за Heers, което позволява на тези растения да се заселят върху кисели, лоши хранителни вещества почви.
Намерен е друг тип - орхидна ендомикороза. Орхидните семена в природата покълват само в присъствието на подходящи гъбички.
Като цяло, в природните условия асоциациите на Mycorris са по-скоро правило, отколкото изключение.
Изследването на останалите съдови растения показва, че ендомикорозата не е била по-рядко срещана с тях, отколкото в съвременните растения. Това даде възможност да се предположи, че появата на микори в процеса на еволюцията може да бъде един от факторите, които позволиха на растенията за уреждане на земята. Като се има предвид слабото развитие на почвата по време на това селище, може да се предположи, че гъбите микориза са в основата на минералното хранене. Съвременните Pioneer растения с микороид имат повече шансове за оцеляване върху бедни почви.
Растенията с микотрофен тип захранване са по-устойчиви на наличието на тежки метали в средата манивогенна среда.
Няколко думи за корените на растенията на влажна тропическа гора. По-често от Wobble, корените на обвивката. Корените приличат на дъските, наклонени с горните краища на дървото. Докато ранените корени са очевидни корени, които се появяват на багажника и растат, образуването на диви корени е свързано със странични корени, нарастващи непосредствено под повърхността на почвата. Страничните корени развиват вертикални растения, триъгълни в напречно сечение, които са в непосредствена близост до ствола на дървото.
Дървеното дърво, често е по-трудно от останалата част от дървото от едно и също дърво, и често те са покрити с тънък слой от кора. Съдейки по пръстените на увеличението, видимо на напречното сечение на тези корени, първоначално е нормално, закръглено в кореновата част, но след това те развиват много силно удебеляване от горната страна. Броят на акчидните корени на дървото варира от 1 до 10, но в повечето случаи има три или повече. В тропиците образуването на корени в гръбначния стълб е особено забележимо в дърветата, които растат в блатисти гори и на лошо изцедени почви. Често корени в гръбначния стълб се използват за промишлени цели. Така че корените на обвивката, които растат в Индонезия на дървото Koompassia Excelsa, отиват на производството на трапезарни маси.
БЯГСТВОТО
Бягство, като корена, е основният вегетативен орган на растението. Escape изпълнява функцията на въздушната енергия, може също така да изпълнява редица други функции и е способна на метаморфоза.
Вегетативно невидогенно бягство се състои от аксиалната част - ствол; Листа и бъбреци. Основната външна функция, разграничаването на бягството от корена, е леденото бягство.
Парцел, който е свързан с появата на странични органи (клони, листа и др.) - възел. Парцел между съседни възли - intezion. Вътрешният ъгъл между листа и стъблото - obsoma Leaf. Всеки повтарящ се елемент на бягство (възел с лист и бъбрек в синуса на листа и интерстициалните) форми Метал. Едва, следователно се състои от серия от метамери, т.е. Тя има метласт.
В началото на 90-те години на 20-ти век американски учени вместо понятията за събранието и междинните междинни анализи за анализ на растежа и диференциацията на растението, концепцията е предложена - елемент на стеблото. Стъбният елемент е част от стъблото между краищата на листния приоритет. Тя се разкрива на издънка от бягство, където няма междукумент. Тези. Новата концепция за "стволов елемент" съответства на концепцията за "метална".
Структурата на апикалната меристем е по-трудна, отколкото в корена. Всъщност, в бягство, апикалната мериста не само образува клетки на първични тъкани, но и участва в образуването на прогресивни листа и упорити бъбреци, които след това се развиват в странични издънки. Това meristem не е защитено от структура, подобна на основния случай.
Видовете Appexes са същите като в корена: монониален, зони, дрямка. Броят на нива е по-голям, отколкото в корена.
Еволюционен примитивен монониален връх. Пример за този тип Appex в коне. Една голяма опъната апикална клетка силно вакубира. Тя е разделена на посоки, успоредна на повърхностите си (с изключение на външната повърхност). Най-близките производни също са значително вакубилирани, но тъй като те са разделени, малките клетки възникват с плътни протопласти. Такива клетки се откриват по периферията на горната част на стъблото, където са положени листите.
Вегетативния апекс бягство на по-голямата част от цветята е изграден по вид туника - корпуси - две зони, характеризиращи се с равнини за клетъчни делении. Туника(Лат. - Shell) включва външни слоеве клетки, които са антикласически (перпендикулярни на повърхността) и се осигурява увеличаване на повърхността. Жилище (Лат. - Тялото) - Централна група от клетки под туниката е разделена на различни равнини, образувайки по-голямата част от бягството. Жилищата и всеки слой туники имат свой собствен първоначал. В туника първоначалното се поставя в центъра по оста. Двойното крайбрежие имат предимно двуслойна туника, едностайна - еднослойна, по-рядко двуслойна.
Докато концепцията за туника - жилището допринася за нашето разбиране за местоположението и растежа на клетките в най-апикалната меристем, разпознаването на различни клетки на клетки в районите в непосредствена близост до него помага да се идентифицира приноса на режимите в диференциацията на тъкани и тела в бягство. Присъствието в апикалите на зони с различни цитологични характеристики е цитохистологична зонилност.
Жилището съответства на клетъчната зона, наречена зона на централни майчини клетки. Клетките на тази зона са силно вакубили - чертата, свързана с относително ниска скорост на митотична активност. Зоната на централните майчински клетки е заобиколена от периферно умение, което се случва в част от туниката, частично от случая. От външния слой на туниката произхожда хистогените на протодермата. Периферната меристем идва от странични производни на апикални инициативи и от централни майчини клетки. Основната меристем се формира в резултат на подразделенията на периферията на централните майки в слоя, наречена преходна зона. Клетките на периферната зона имат дебели протопласти, следователно имат висока митотична активност.
На определени места се поставят листните туберкули (или първични туберкули) с особено висока активност.
Появата на нов лист приоритет е свързан с промяна в посоката на клетъчни дивиденти, която е предшествана от промяна в ориентацията на микротубулите в периферните слоеве на цитоплазмата. Осмотичното налягане влияе и върху растежа на приоритет.
През периода на активен растеж апикалната мериза на бягство образува листни страни толкова бързо, че възлите и интерстините не са различни в началото. Постепенно, местата между листата на листата започват да растат, разтягат и придобиват вида на близките. В същото време зоната за свързване на листа е изтеглена в възли.
По-долу, когато се появява такелаж на листата, се наблюдава вакуолин. Това показва началото на диференциацията на първичната кора и ядрото. Силният вакуема в първичната кора и ядрото показва наличието на мерматична зона, наречена остатъчна меристи. Изглежда, ако има апикален остатък, запазен сред по-диференцирани тъкани. По-долу е мястото на листовото фокусиране в остатъчната меристи чрез надлъжни разделения, без последващия растеж на клетките в ширина, се образуват няколко удължени клетки на първата принонбия.
На следващите етапи на развитие в остатъчната меристем все по-често изпомпващите ленти се диференцират. След като бяха оформени всички провеждащи снопове на това ниво на стъблото, остатъчната меристем се диференцира във взаимното разделяне на паренхима. В възлите, част от остатъчната meristem става паренлив слот за листа.
Provinbium може да бъде фиксиран с твърд пръстен или отделни тежки овен. Ако Приронбини лежи с твърд пръстен, образуван apple (неизпълнен) вид структура на стъблото. Ако водопроводът е положен поотделно, той се образува пъзел тип стъбло. Следователно, в стъблото, първичните водещи тъкани се поставят по различни начини: в първия случай, твърд пръстен, във втория - отделни проводящи лъчи, разделени от основните паренхимни лъчи.
В обезпечения проводящи греди, първичната флора се появява във външната част на помпената лента, първичния ксислам - във вътрешния. Последваща диференциация на етажа се извършва центропетал: новите елементи на Flolam изглеждат по-близо до центъра на стъблото. Ксилем се диференцира в обратна посока - центробежен.
Цялата стела, или централната част на стъблото се състои от проводими тъкани, ядра (понякога се срутват), периколеси. Началната кора е разположена извън перикола. В състава на първичната кора (и произвежда техните периферни меристи), включва хлорохенхем, понякога секреторни елементи. Най-външният слой на периферните меристи не образува екзодерма, както в корена и се диференцира в колежа, което се крие или под формата на тежка материя или твърд пръстен (в зависимост от вида на структурата на стъблото: лъч или глупост). Вътрешният слой на първичната кора - ендодерма. Но в стеблото, той не извършва регулаторната функция, както в корена. Зърната на нишестите са положени в ендодерм и затова се нарича скорбяла влагалището. Понякога има калциеви оксалатни кристали.
Персичкуването бързо губи по меристематичната активност и се превръща в склеренхим, който се среща с твърд слой или тежък, в зависимост от вида на структурата на стъблото.
Ядрото е представено от тънкостен паренхимни клетки, в които се отлагат резервни хранителни вещества. Има клетки - идиосласти, пълни с кристали, слуз и др. Периферната част на ядрото се нарича perimedullar Zone. (Гръцки. Пери - около, медула - ядро). Клетките на перимедлулалната зона са по-малки, дълго запазват препитанието си.
Източникът на монокотелинг е вид лъч на структурата на стъблото, т.е. Provinbium се намира с тежки. Но вторичната меристема - Cambier - никога не се случва и следователно няма вторични проводими тъкани. Дори и с удебеляване, стъблото се образува само от първични тъкани. Дебелото монокуларно стъбло (Palm, Aloe) се развива поради разделянето и разтягането на клетките на главния паренхим. Този растеж се нарича дифузен вторичен растеж.
Моноцикъл Първичната кора и централният цилиндър не са разделени от ясна граница, като два класа и Stele се приближава към епидермиса.
За една спалня, дифузният тип на структурата на стеблото е характерно, когато затворените проводящи лъчи са разпръснати по време на дръжката. В процеса на удължаване на интерстициал, първичните елементи на FLEEM и Xylems (протофлоем и протокол) са разтегнати и унищожени, в резултат на което се образува голяма въздушна кухина от ксилемата на лъча. Ето защо, в лабораторни упражнения, когато наблюдаваме видовете греди, на постоянно напречно рязане на царевичното стъбло, беше ясно, че зрелия проводящ лъч включва два широки съдове на метаксале, въздушна кухина, метафлома. Всеки пакет е затворен в затягане от склероронс.
Монокуните често умират ядрото на стъблото и кухината се оформя в нейното място. Така се нарича такова кухо стъбло с добре изразени възли соломина. Солмин се характеризира със силно развитие на механичната тъкан, която образува пръстен върху периферията на стъблото. В зрели растения хлориним се унищожава, а Солкин придобива златен жълт цвят, характерен за склеренхим. Като цяло, монокуларната характеристика се характеризира със силна стебла, когато до 30% от всички тъкани попадат върху Склероштин и може да има частично лигнифициране на свързани паренхимни клетки.
В стъблото на дикотислоен вторичен растеж е резултат от Камбия. Cambier се появява в края на първата година на растежа на бягството и допринася за удебеляване на стъблото. Cambier се среща частично от пронамбия в проводящи греди и частично от интертекския паренхим. В зависимост от мястото на образуването, секциите на Камбия се наричат \u200b\u200bпакети и интеркуети камбики. Бъдещите камбиални клетки в интер дяловете се определят много преди началото на камбиална активност. Cambier се образува първо в клетките в непосредствена близост до гредата, след това се инкубира дълбоко в сърцевината. В резултат на това пресичането и лъчът камбийс образуват твърд камбиален пръстен в стеблото.
Разграничават се три вида вторичен растеж:
1) Тип Kirkazon Aristolochia.
Първоначалният тип лъч на структурата на стъблото. Първичните проводими лъчи са разделени от широки лъчи. Гръбният камбий формира вторични проводими тъкани, междубукан-радиационен паренхим. Такава пакетна структура се запазва.
2) Helianthus слънчоглед тип.
Също така, първоначалният тип лъч е вид структура. Но вторичните провеждащи тъкани се формират както на лъч, така и междукубни камбики. Вторичните проводими греди, образувани от interochetic камбия, се различават от първични по-малки размери и отсъствие върху куп механична тъкан перицикличен произход. При продължително действие на Cambia, вторичните греди стават все повече и повече. Те растат, обединяват се помежду си, образуват солиден пръстен от вторични проводими тъкани.
3) тип Tilia Lipa.
Първоначалният е видът на структурата на стъблото. Общият камбиален пръстен образува вторични проводими тъкани. Но от време на време Камбий образува паренхимни клетки, които са допълнително диференцирани във вторичните лъчи.
Структура на напреженията на стъблото
Дървесина и храсталаци, както и иглолистни, вторично удебеляване продължават в продължение на много години, а стволовете в някои видове достигат няколко метра в диаметър. Удебелението се свързва с дейностите на дълга функционираща кампания и частично гълъб - две средни служби. Топографски в стеблото на многогодишно дърво растение може да се отличава с три основни части: кора, дърво и сърцевина. Границата на кората и дървото преминава през Камбия.
Cambia клетки, за разлика от типичните мерматични клетки, винаги са много вакубили. Състоянието на Cambia клетки Цитоплазма се променя през сезоните.
В камбиалната зона или камбиалния пръстен има две форми на камбия клетки: удължени клетки с форма на гръбначния стълб (първоначално) и удължени сферични клетки - радиален първоначал в хоризонтално посоката. Ако клетките с форма на шпиндела са поставени подходящи хоризонтални редове, цемът се наричат \u200b\u200bдълго. Ако няма такъв ред при поставянето на предевоидни клетки - не-твърди камбий.
Liewer Cambier дава началото на дълготрайна дървесина и дългосрочен единен крем. От не-русоларната камбия произлизат не-русово дърво и небималична флора. В еволюционния план дългоразмерените фло се считат за по-високо специализирани от не-библиотеката.
С удебеляване на стъблото, обиколката на слоевете на Камбия се увеличава. Това е възможно поради разделянето на камбиалните клетки в радиалната посока.
Сред камбиалните клетки се отличават със средния първоначален слой. Клетките от него в резултат на деривативи от разделяне във вътрешния, и след това във външната посока. За същия период от време се отлагат 2-3 клетки, а във външната посока е само една клетка. В резултат на дърво се образува повече от луба.
Радиационни клетки (начални) образуват основни панеримни лъчи: първичен, който се разтяга от ядрото към кора; и вторичните лъчи са по-къси, защото възникват от радиални клетки, които са възникнали по-късно.
Групите могат да бъдат едноредови и множествени; хомогенна (т.е. състояща се от идентични клетки) и хетерогенна (съставът на лъча включва клетки, различни в структурата).
Стойността на сърцевите лъчи : Това е мястото на снабдяване с хранителни вещества (нишесте, липиди), която играе роля в периода на почивка (например през зимата).
Гръдните лъчи осигуряват радиален трансфер на вода и минерални соли от ксилари по апопласт до камбия и вторична флора и в обратна посока по симпелатите на хранителните вещества.
И накрая, обменът на газ се извършва върху интерклодерите.
дърво
Дървото е оформено на Камбие и се увеличава в центробежната посока. Като В нашата зона камбий произвежда периодично: той започва да функционира рано през пролетта и спира активността в края на лятото, концентричното наслояване се вижда в дървото. През пролетта в началото на вегетационния сезон се формира ранното дърво. Неговите кораби са широки, тънки стени, способни да извършват значително количество вода. Водата е необходима за иницииране на растежа, особено за увеличаване на размерите на нови клетки (например клетки на развиващи се листа). Късно дърво съдържа по-малко съдове. Те са по-тесни, дебели оградени. Преходът от по-рано до късната дървесина от същата година може да бъде постепенно. Въпреки това, границата на късната дървесина от една година с ранната дървесина на следващата година винаги е остра и ясно видима на напречното сечение. В резултат на това се образува годишно увеличение (търговски пръстен или едногодишен слой).
При поставянето на кораби в годишното (годишно) се увеличават следните видове дървесина.
Ако съдовете са разположени главно в ранна дървесина, а по-късно те липсват или са поставени само малки плавателни съдове, като дъбова дървесина, пепел, е Callingesewood Wood..
Ако съдовете са разположени равномерно в годишната печалба, въпреки че диаметърът им в късната дървесина намалява, е Разпръснато дърво. Намира се на бреза, клен, топола.
Има преходни форми между тези видове дървесина.
Тъй като дървото изпълнява проводими, механични и отгледи функции, то включва механичната кърпа под формата на склеренхим и под формата на основния паренхим. Структурата на всички анатомични елементи, техният брой, поставянето дава на дървета с различни растения специфични характеристики, характерни за конкретна таксономична група.
Например, специализирана механична тъкан (Sclerchima) има специализирана механична тъкан. Примитивен в еволюционен план. Ди-дебелото черво (магнолия) нямат дървени влакна. Механичната роля в този случай се извършва от трахеди.
В дърветата на някои растения има преходни форми от трахед към Libraform (либриформи - специализирани механични елементи на дърво, състоящи се от прокураст, насочени към краищата на клетките с дебели изветрени черупки). Възникват разделени библиоформи. Първоначалната клетка с форма на ремък с дебели вертикални черупки е разделена на отделни къси живи клетки. Такива хистологични елементи са морфологично и функционално приближават дървения паренхим.
Дървевец Parenchyma изпълнява здрава и частично проводима роля. Клетки паренхим живи. Поставени дървени паренхимни хоризонтални и вертикални редове. Хоризонтални редове образуват сърцевини. Техните функции, които вече сме разглеждали. Вертикалните редове образуват самия горист паренхим.
Ако дървесният паренхим е разпръснат през годишното увеличение, то Дифузен паренхима. Характеристика за дървената липа, дъб, круши.
Ако дървесният паренхим се поставя около съдовете, то паратрахеален паренхим. В гората на пепел, клен.
На външната граница на годишния растеж се намира терминал Паренхим. Пример: дърво върба, лиственица, магнолия.
Особеността на неговата структура е липсата на кораби. Трахелските елементи са неработещи и представлявани от трахеди, които извършват и проводим и механична роля.
За трахед иглолистни дървета се характеризират с големи пори с Torr. Трахедс са дълги с заострени краища. Ранните трахеди са големи, тънки стени. Късните трахеди са сплескани в радиалната посока, дебел стени.
Виждаме, че дървото на гласовете и дикотилдоните растения имат значителни структурни различия. Ето защо тя е разделена на две основни групи. Дървените подаръци са меки и дървото на дикотомично - към масивни гори. Тези термини "твърди" и "меки" не са непременно намалени само до плътност и твърдост. Разликите са свързани, както вече казахме, със структурни характеристики.
В наклонени или извити стволове се появява реактивна дървесина. Неговото образование е причинено от тенденцията на клонове и стволове, за да се противопостави на товари, произтичащи от наклонената позиция, т.е. Неговото образование е свързано с процеса на изправяне на тези части на растението.
Най-важните фактори, причинени от развитието на струйно дърво, са гравитационни сили и дистрибуция на ендогенни стимуланти на растеж.
Реактивната дървесина е различна от обичайната и анатомично и химически. Клетките на струйната дървесина са подложени на по-голяма лигнификация и удебеляване на обвивката. На иглолистни е по-плътна и тъмна от заобикалящата тъкан. Нейните трахеди са по-къси от трахед на нормалното дърво.
Радиоактивността на стеблото намалява от външните слоеве към ядрото. Има известно увеличение на замърсяването на дървото от основата на багажника до върха.
Достатъчно високо съдържание на радиоактивни вещества в кора и издънки показва тяхното външно въздушно замърсяване.
Лай
Този термин съчетава всички тъкани извън Камбия. Това вече е вторична кора, защото Той се формира от вторичното измерване - камбий (за разлика от първичното, което се оформя по време на диференциацията на клетките на първичната меристи).
Вторичната кора включва вторична флора, sclernerehim (Luban влакна и каменисти клетки), основната паренхима. Нарича се комбинацията от механични елементи твърд лоб Проводими елементи и основен паренхим - мек лоб.
В вторичната корка има секреторни клетки, канали за смола. Основната функция на вторичния кортекс е поведението на пластмасови вещества, също така функцията на защита.
Вторичният етаж представлява значително по-малък дял на обема на цевта, отколкото на вторичния ксислам. Това се дължи на факта, че първо, Камбий образува по-малко флора от ксиславе и, второ, старото, не функциониращо платно, постепенно се смачка.
Благодарение на увеличаването на площта на аксиалния орган, паренхимните лъчи се разширяват и приличат на триъгълник, чийто връх е насочен към Камбия. Такъв паренхим се нарича дилатация (lat.dilatatio - разширяване). Такова поставяне на паренхима защитава лоб от прекъсването, когато стъблото в дебелината се увеличава.
В иглолистните, вторичната флора има по-проста структура. В луббените хистологични елементи се поставят равномерно и, разбира се, присъстват смолни канали.
Всички тъкани извън Meltogen (означават най-вътрешният корк камбий), тъй като клетките на корк са отделени, вода и минерални вещества се прекратяват. Всички тези перидмс с останките на кората (фиктивни клетки на паренхима и етаж) съставляват външна (външна) кора. Живата част на кората между Камбие и вътрешният слой на гълъба се нарича вътрешна кора.
Така че, разчитайки на нашите познания от темата "тъкани", погледнахме анатомичната структура на стъблото като аксиално бягство.
Сега помислете за структурата на страничната организация на бягството.
ЛИСТ
Лист е основният фотосинтетичен орган на висшите растения. Структурата на листа и нейните функции са тясно свързани.
От общото уравнение на реакциите на фотосинтезата:
CO 2 + H2OH → HV (CH2O) N + O2
може да се заключи, че 1) листата се нуждаят от източник на CO 2 и вода; 2) листата трябва да бъдат адаптирани да абсорбират слънчева енергияте трябва да имат хлорофил; 3) като един от реакционните продукти ще се открои около 2; 4) въглехидрати трябва или се отлагат или транспортират до други части на растението. Лист е много специализирано тяло, което отговаря на всички тези изисквания.
Цялата еволюция на листа, като вегетативен орган, се проведе по пътя на развитието на приспособимостта най-добра употреба Света.
Според микрофиловата линия на еволюцията, листът се счита за архаични растения от вида на ринове, който запазва структурата на аксиалната структура на органите.
Според макрофилната линия на еволюцията, листото възниква в резултат на изравняване, поставянето на тялото в една равнина и последващата комбинация от тях в едно. В същото време, способността за дългосрочно потвърждаване и разклоняване е загубена. Какви са морфологичните и анатомични характеристики на лист, произтичащ от дълга еволюция?
Подходящ лист обикновено се състои от листова плоча и домашен любимец. Черкок - тясна основна част на листа между листата и бутона за бягство, с който листът е фокусиран в пространството и се публикува най-благоприятно за светлината. Чрез пълнежа е връзката на листната плоча и стеблото. Режещите и механичните тъкани са добре развити в разрез. С листа попада в разрез, се образува отделящ слой.
Нарича се най-нисък лист, съчленен със стъблото основата на листа. Базата на листа може да се вземе разнообразна форма. Най-често има формата на малко удебеляване и наречено листова подложка. По-често на базата на листа има различни форми и размер на сдвоените странични растеж - домакини. Акцентите могат да бъдат листни и да служат като допълнителна асимилираща повърхност. Акценти могат да бъдат необходими и да извършват функции за защита на растенията.
Има форма с надраскване, филмови стриптизьорки. В еволюционния план има намаляване на заварчиците и еволюционно млади растения, те изобщо не са рано.
В някои зърнени култури, целина (чадър), основата на листа създава и образува затворена или отключена тръба - листа вагина. Листата вагина допринася за дългосрочното запазване на междукалаторните меристи и служи като средство за допълнителна подкрепа за бягство.
Основната част от листа е листа. В резултат на неравномерно осветяване на горната и долната страна на плочата на плочата има атропентрална структура, т.е. Структурата на горната страна на листа, коремната, се различава от структурата на долната страна, гръбната част. Горната страна на листа се нарича коремна, защото В бъбреците тази страна е вътрешна и се превръща в осната ос. Колкото по-нисък, гръб, страната на листа в бъбреците е обърната патица от растението. По този начин, листа с анорентрелна симетрия е двустранно или пулсиално или св. Характеризиращ се с дискриминално.
Лист, имащ радиална симетрия, е уравнителен. Такива удебелени, цилиндрични листа се намират в Толстанков. Едностранно или изолатерално унифициално, листата са характерни за зърнените култури, лук.
Обмисли Анатомична структура на листова плоча.
Подобно на корен и стъбло, листът се състои от покритие, проводима и основна тъкан. Като Листът обикновено няма вторичен растеж (с изключение на незначителен растеж в твърд и големи вени), епидердата се запазва като покриваща тъкан.
Ustiana се намира главно на долната повърхност на листа. В листата на дикотилиран решетка прах, разпръснати по цялата повърхност на листа, няма очевидно ред. В едностайни и иглолистни листа те се разпространяват от редове, успоредни на надлъжната ос на листа. В епидерм на някои зърнени култури има моторни или моторни клетки. Те са по-големи от обикновените епидермични клетки. Вакуорът заема почти цялата клетка. Със загубата на тургората тези клетки се намаляват и помагат за сгъване или завъртане на листа.
Повечето от Основната тъкан на листната плоча се отчита от мезофил. Мезофил диференциран. Palisada Mesophyll се поставя в горната страна на плочата, гъба - на дъното. Листата с такава структура и има сръбски или дълбоко.
Ако панелът мезофил е поставен от двете страни на листа, листът е дезинфектен или изолатерален.
В мезофил на зърнени култури на различна диференциация на парализата и гъба хлорохенхем. Мезофилите клетки се намират радиално около гредите.
Проводими листови снопове се наричат \u200b\u200bжители и разклоняващата система на жителите - място. Характеристиките на местоположението на проводящите греди в листните плочи определят различни видове Жилищни, които могат да бъдат дихотомни, паралелни, аркиидни, палпа и купчина.
В зависимост от вените на по-високи крайни поръчки са свързани помежду си, разграничават отворените (дихотомни) и затворени корпус. С дихотомното корпус, алкалният клон дихотомично се отклонявайте под остър ъгъл и стигнете до ръба на листата, без да се сливате със съседни вени. Със затворен корпус, малки вени в резултат на анастази (интерстигации) образуват мрежа, която е проникнала с цялата плоча или по-голямата част от нея.
Листата с пръст и цигари, и това е най-вече храносмилателно, имайте средата, най-голямата вена. Вената се състои от първичен ксислам и първични плавания, произтичащи от Принонбия и се съчетават в обезпечения лъчи. Камбият често се образува между ксилем и платието, но не работи. Следователно, xilem и flym са първични.
Бучките образуват непрекъсната система в лист, свързан с проводимата система на стеблото. Следователно, Xylem винаги е фокусиран върху морфологично горната страна на листа, а платието е морфологично по-нисък.
Средният воал е свързан странично с величината на страничните линии. Всеки от тях е свързан с още по-малък, а тези от своя страна са разчленени за още по-малки и т.н. Броят на разклоненията на разклонението варира от 2 до 5 или повече. Най-малките форми образуват клетки, в които са затворени малки площи от мезофил - ареола.
В отворена дихотомна система, отсъстват затворените клетки.
В едностайни листа (с паралелен корпус), надлъжните вени са взаимосвързани с по-малки вени - пунктирани греди, които са подредени под формата на прости напречни джъмпери. Количеството на тъканите проводими в малки вени постепенно намалява. В края на гредите, ксиловите елементи често се изтеглят на по-голямо разстояние от phloem. Ксилем в края на вените обикновено се състои от къси трахеални елементи, платието е от къси тесни сито и широкомащабни сателитни клетки.
Малки провеждащи пакети, локализирани в мезофил, са заобиколени от един или повече слоеве от компактно сгънати клетки, образуващи скоба с лъч. Punch Clamps може да бъде паренхим или склероронс. При някои видове има суберизация на клетъчните мембрани в плаките на гредите, което показва, че покритите клетки могат да функционират като ендодерма.
В големи вени с една или от двете страни има колеж и склерошим. Тъканта, свързана с големи вена, е кули над повърхността на листа и образуват издатини. Парцел, сключен между издатините на вените, се нарича "интеркостална зона" (лат. Името на издатината "Коста" е ръб).
При видове с дебели листа, в мезофил има различни склейдс, които извършват характеристиката на подпора.
Листата от зърнени култури се характеризират със силно развитие на склеренхим.
Листата на гамоторите са по-малко разнообразни в структурата, отколкото листата на покрития мост. Помислете за примера на шегувия на бор, като годността за прехвърляне на ниски температури през зимата и способността за живот в продължение на няколко години е отразена в анатомичната структура. Свързването има дебелостенна епидерма с мощна кутикула и дълбоко потопени астейци. Някои иглолистни клетки (cupressaceae) имат козина ролка, която виси над затворените клетки на UST. Stwitz се поставят от вертикални редове от всички страни на chevyo. Под епидермма има дебела стена с частично от лигираните черупки на клетките на хиподермата. Мезофил сгънат. Той съдържа най-малки движения. В центъра на шевните са проводими греди. Обикновено две, по-малко от един. Тези греди са свързани с склеренхимна замазка. Греймите са заобиколени от трансферична тъкан, състояща се от трахеид и паренхимни клетки. Трансфузионната тъкан (LAT. Transfusio - трансфузия) е свързана с транспортирането на вода и хранителни вещества между проводящите греди и мезофил.
Тъканната трансфузионна тъкан е заобиколена от дебела оградена ендодерма. Endoderma често разполага с капкова почивка в ранните етапи на развитие и суберската плоча по-късно. В зрялото състояние на ендодермените клетки имат вторични линевни черупки.
Анатомичната структура на листната плоча разкрива значителна пластичност. Структурата варира в зависимост от условията на растението и поставянето на листата на растението.
Вече отбелязахме една от указанията за развитие на съвременната ботаника - екологичната анатомия на растенията - и каза, че животът на килията - протопласт - главно реагира на промяна в условията на околната среда. Така че клетките на северните растения са по-малко вакубилизирани, те имат големи хлоропласти. В строма се откриват много нишестени зърна. Тилакоидната система на хлоропластите е по-слаба, отколкото в хлоропластите на растенията с умерени ширини. Броят на митохондриите в северните растения е 2-2.5 пъти по-висок. EPR и устройството Golgji са по-силни. Такива особености на субмоскопската структура на мезофилните клетки на растението на север се характеризират като важни адаптивни знаци, които допринасят за оцеляването на растенията в сурови климатични условия.
При растенията, адаптирани към дефицит на влага, се празнуват студени клетъчни мембрани, особено в епидерм. Силно удебелени външни черупки на големи клетъчни епидермиса заемат половината от кухината на клетката. Над епидерматума е дебел слой на кутикула. От повърхността на кутикулата целият восък е покрит с дебели скали, от които зависи от листата на листата.
Прахът е потопен в вдлъбнатините, изпратени от епидермални косми.
Сочни растения имат наличието на вода за водоснабдяване.
Интересни устройства за улавяне на влага в листата на лебед Софт (Африка). Листът от горната и долната страна е покрит с твърд слой от множество поставени мехурчета. Дебелината на този слой значително надвишава дебелината на мезофила и предпазва живите тъкани от сушене. Косата могат да хванат въздушната влага през нощта и да подобрят суровия воден режим на растението.
Листата и стъблото на тампоните на кодираните яйца на нашата горска зона са покрити с множество балонни косми, които съставляват впечатлението за измъчена плака, но такъв гъст многоредов слой, като мек лебед, те не се образуват .
Използвайки примера за сравняване на листата на два вида лебедки, може да се види, че системните признаци на близки таксономични единици в презаселването им в различни условия на околната среда придобиват неравностойно развитие и са адаптивни.
Листата от тропически гори (например чудовище) имат добре дефинирани листни плочи и дълги скокове. Излишната вода винаги е лесна за работа. Ustian Open. Палусадична тъкан с големи въздушни кухини.
В допълнение към температурата, влажността е друг важен абиотичен фактор - светлината, върху която растението реагира със структурни промени.
Листа, развиващи се в пряка слънчева светлина, по-малка сянка, но по-дебел от сянка в резултат на силното развитие на пелеозен мезофил.
Така, залегнало по време на развитието на адаптацията към условията на средата, се отнасят до количествени различия, но не променят принципите на организацията.
Едногогенезен лист
Листата се полагат под формата на малки туберкули като страничен растеж на апикалната меристи. Не само туника, но и клетъчни клетки участват в образуването на листната туберчана. Изброеният приоритет нараства от първоначалната издатина под формата на конична или игла. Скоро листата са заобиколени от апик на апикал, предпазват го и механично, и тези топлина, които са изолирани по време на дишането.
Растежният растеж на листа обикновено се разделя на апикален и маргинален (ръб). Първият е свързан с удължаването на приоритета, вторият - със страничен растеж, в резултат на което се образуват две части от плочата. В съответствие с това нарастващият приоритет има апикална мериста на върха и две маргинални меристеми от двете страни на оста. Апичният растеж на Прим не е дълъг. Растежът на върха и краищата на записа е растеж на междувалята.
В сложен лист активността на маргиналната меристем се локализирана в отделни центрове, всяка от които образува отделна листовка.
Развитието на листата е по-бързо от височината на дръжката, която се формира по-късно от междукаланския растеж.
Ако на границата между базата на интеркаланските и маргиналните зони се образува теглене, се развива лист за възглавница. По-голямата част от дръжката се формира от междукалаторната зона. Ако такъв влекач (на границата между базата на интеркаланските и маргиналните зони) не се формира, се развива заседнал лист.
В листа на последователността на монокотильозните растения, пределните меристеми почти не са положени и дълго време се характеризира с междукаленски растеж. Оттук и основната форма на линейна плоча - линейна.
Разликите в степента на разделения и разтягащите клетки от различни слоеве на плочите водят до образуването на многобройни взаимодействия и типичната структура на мезофил.
Развитието на други тъкани корелира с развитието на проводящите тъкани. PROVINBIUM се изолира едновременно в листа и съседна част от конуса на увеличаването на конуса, образувайки непрекъснат съдебен процес. Той образува първичен проводящ лъч, който е непрекъснат и общ за лист и ствол.
Онтегенезата на листа показва общата природа на листа и стъблото като част от цялото - бягство.
Тъй като разгръщането на бъбреците започва абстрактната фаза на развитието на листата. Повърхността на листата се увеличава след няколкостотин, дори хиляди пъти. Нарастването на повърхността се постига чрез разделяне на повечето листови клетки и ги разтега по дължина и ширина.
Продължителността на живота на листата зависи от генетичните и климатичните фактори. В решаващи дървета и храсти на умерен климат, покрайнините на листата са 4-5 месеца. Листата от иглолистни растения живеят от две до шест или повече години. Но във всички случаи продължителността на живота на листата е значително по-малка от продължителността на живота на аксиалните органи на растението.
Welwitschia Mirabilis расте в скалисти пустини в Югозападна Африка. Това дърво - джудже има дълъг корен, дебел и къс багажник (висок до 50 см и с дебелина до 1 м) и два големи листа, които остават през целия живот (до 2000 години). Листата достигат 2-3 метра дължина и постоянно растат в основата, движещи се в върховете. Почти единственият източник на влага за Велчичия е гъста мъгла, чиято влага това растение абсорбира през многобройния прах от двете страни на листа (22 200 праха на 1 cm 2).
Активното разделяне на листата от клоните без увреждане на живите му тъкани се нарича обжалване. Сезонният спад на дърветата е резултат от реакция на промяна в дължината на деня. Изплащането на листата в есента значително намалява повърхността на изпаряването на растението, което е необходимо при условията на физиологичен дефицит на влага през есента и зимата. Благодарение на падането на листата, опасността от половината от клоните от тежестта на снега.
В повечето листа, образуването на слой на разделяне възниква по време на онтогенезата. В тази зона количеството механична тъкан се намалява. Плочките се образуват в трахеалните елементи, преди да попаднете в трахеалните елементи, Callose се отлага в ситото елементи. По време на освобождаването ензимното разрушаване на клетъчните измервателни уреди, което води до отделяне на клетките един от друг. Промените в черупките включват загуба на циментираща способност на средната плоча (частично поради отстраняването на калций от него), хидролизата на самите целулозни обвивки и пролуката на увеличените трахеални елементи.
Защитният слой или белег, се оформят в резултат на отлагания в черупки и взаимодействия на защитни вещества, като суберин или рана. В дървените растения защитният слой се заменя с перидермата, която е поставена под защитен слой И става непрекъснато с перидермата на останалата част от стъблото.
Падането на листа не е непременно свързано с процеса на разтваряне на клетъчни измервателни уреди. Когато отпада иглите, може да се наблюдава механично разделяне на без предишни химически промени.
След привлекателността на листата, листните белези остават с белезите си. Лъч белези - Това са счупени краища на проводими греди, които вървяха от листните следи в лист лист към жалбата си.
Листови следи - Това е клон от проводимата система на стъблото на листата. Листата се простира от мястото на сливането му с ударния лъч, докато листът бъде освободен. Един лист може да има една или повече следи от листа.
Като Стъблото и листът имат филогенетично общ произход, без фундаментална разлика между листовите следи и лъчите на стъблото. Съответните термини имат описателна, топографска стойност.
В зоната на възела, където листовата пътека се отклонява от централния проводящ цилиндър на стъблото в посоката към основата на листа, паренхимна зона се оформя в цилиндъра - листа лакуна или слот за листа. Броят на листовите следи и листата лакуна е различен в различни растения.
Основният тип възел в покрития мост е възел с три или повече лакуна. Основната посока на еволюцията на покрития мост се характеризира с намаление на количеството пресъхват.
Lacumba, сърцевински, междуфункционални зони, проводими и разположени на периферията на проводящата система Pericycle съставляват вътрешния прът на аксиалната част на растението (стъбло и корен) или стомана.
Класификацията на Stelle се основава на взаимното подреждане на проводими и непроводими тъкани в аксиалните органи на първичния етап на развитие. В най-простия тип стел, който се счита за най-филогенетично примитивна, проводящата тъкан образува твърда колона и платието заобикаля ксилем. Това беше протестично. Такава организация на стъблото е биологично несъвършена, защото Повърхността на контакт между провеждащата и основните тъкани е малка.
По-нататъшното еволюция на Stelle беше на път за увеличаване на повърхността на контакта на продощи тъканите с главната. Това беше постигнато по два начина. В един случай, Stela образува дълбоко отглеждане отстрани на кора и на кръста наряза има форма на звезда. Това е актьотел.
Вторият, биологично по-обещаващ път на развитие е появата на паренхимно ядро \u200b\u200bв центъра на Стела. Това е синологична.
По-тясна връзка между проводимите и основните тъкани възниква с развитието на листата. От него още листаОсвен това, Stele е наситена с основния паренхим. Stela придобива вида на мрежата (гръцки. Network - dictuon). Беше актьор.
Още по-паренхимализиран стил от модерно цвете. Stela, която е система от проводяща течение и пресичащи се зони, - усел (гръцки. ЕС е добър).
Листовите следи от одрачос достигат до центъра на стъблото, след това отклоняват периферията. Ето защо, на напречното сечение на стъблото, гредите изглеждат разпръснати без никаква поръчка. Атактос с гръцки. разхвърлян. Оттук и името на Singlean Stelny - Attaketosla. Характеризира се с висока степен на паренхимализация, когато всеки листов отпечатък & quot



































- Размер: 2 mgabytes
- Брой слайдове: 36
Описание на презентационния корен и коренови системи 1. Функции и еволюционни слайдове
Коренови и коренови системи 1. Функции и еволюционен корен. 2. основната структура на корена. 3. Вторични промени на кората. 4. образуването на странични и очевидни корени. Коренови системи. 5. Специализация и модификации на корените.
Коренът е аксиален орган с радиална симетрия и неясно все по-често се дължи на дейността на апикалната мериста. В корена листата никога не се появяват, а апикалната меристем винаги е покрита с пещерня. Основната функция на корена е абсорбцията на вода и минерални вещества, т.е. осигуряване на почво хранене на растението. В допълнение към посочената основна функция, корените се изпълняват от други функции: укрепване на растението в почвата, дават възможност за вертикален растеж и стрелба; Корените се срещат вторичният синтез на различни вещества (аминокиселини, алкалоиди, фетитогоровини и др.); В корените могат да бъдат положени резервни вещества; Корените взаимодействат с корените на други растения, почвени микроорганизми и гъби.
Корените възникват от тялото на ринофит, отворени по повърхността на почвата. В хода на еволюцията някои клонове на тези тела започнаха да се ровят в почвата и да доведат до корените.
Корените са адаптирани към по-съвършено хранене на почвата. Външният вид на корените беше придружен от дълбоко преструктуриране на цялата им структура. Те имаха специализирани тъкани. Функцията на абсорбцията на вещества от почвата започна да извършва млади покрития на корените. Те запазват живи клетки на повърхността. Тези клетки са образували коренната тъкан - risoderma във функционалност.
Функцията на абсорбцията на вещества от почвата започна да извършва млади покрития на корените. Те запазват живи клетки на повърхността. Тези клетки са образували коренната тъкан - risoderma във функционалност. След това в процеса на еволюцията се наблюдава увеличение на абсорбиращата повърхност на корена поради трите фактора: 1) изобилно разклоняване и образуване на голям брой смукателни окончания; 2) постоянно увеличаване на корените и движението на изсмукване на засмукване в нови части на почвата; 3) образуването на коренни косми.
Тъй като възникването на корена възниква в гъста почва, топ меристемата трябва да бъде защитена. Защитата на APICAL MERISTEM от щети гарантира появата на основен случай. Появата на корените е причинена от увеличаване на климатичната сухота. Появата на по-сух климат е причинила наземни растения необходимостта да се прикрепят вода и хранителни вещества на субстрата и да абсорбира от него. Въпреки това, по време на еволюцията, структурата на корена в различни видове растения се е променила по-малко от това на стъблото. Това се дължи на факта, че в почвата условията са по-стабилни, отколкото във въздуха. Следователно, коренът се счита за по-"консервативна" власт, въпреки че изглеждаше много по-късно бягство. Образуване на корените - важна ароморфоза на растенията. Благодарение на него, растенията успяха да научат повече суха почва и да образуват големи, като се издигат издънки.
Реакция на асиндласти от основен случай за гравитация. Движението на статиолитите играе важна роля в създаването на градиенти на фитогорите, осигурявайки вертикален растеж на корена.
Структурата на корена на разсад на пшеницата (Triticum aestivum): диаграма на структурата на корените; B - Диференциране на клетките на ризодермата и екзодерма. 1 е зона на поведение, 2 - зона на засмукване, 3 - зона на разтягане, 4 - зона на дивизия, 5 - коренни косми, 6 - корен.
Кръст на корена (а - моноцикъл, b - велосипед)
Първичната кора се появява от библерса. Неговата основна маса съставлява живите паренхимни клетки с тънки черупки. Между тях се образува системата от взаимовръзки, опънати по протежение на основната ос. Циркулират газове (CO 2) за интерклодерите. Газовете са необходими за поддържане на интензивен метаболизъм в клетките на кората и ризодермата. Енергичният метаболизъм в клетките на кората е необходим за изпълнение на редица важни функции: 1) кортекс клетки захранват с ризодерма с пластмасови вещества и участват в абсорбцията и провеждането на вещества; 2) в основата се синтезират различни вещества, предавани на други тъкани; 3) резервни вещества се натрупват в кората клетки; 4) Често има GIF файлове на гъби, образуващи Mikuriza.
Клетките на Endoderma преминават три етапа на развитие. В смукателната зона ендодермата е на първия етап. В средата на радиалните стени на клетките му се образуват каппорните колани. Коланите на Каспари се припокриват с движението на вещества през клетъчните черупки, т.е. според Apoplast. Вторият етап може да се наблюдава в страничната корена. В същото време S. атрешна Клетъчната обвивка се появява тънка шипка. Въпреки това, endoderma все още е свободен да преминава решения, тъй като остава отделни пропуски с тънки стени. Третият етап от морския бряг и ръба на ендодермата може да се наблюдава в зоната на монооценности. Вътрешните и радиалните стени на клетките му са много удебелени. На напречни секции, такива клетки имат подкова с форма на подкова. Няма клетки на честотната лента. Tolstyed endoderma защитава проводящите тъкани и увеличава силата на корена.
Каспари - водоустойчива бариера, принуждавайки вода да остави апопластичната и да бърза през мембраните на клетките на ендодермата в симпласт
Потокът на водата от почвата към корена: водата може да се движи около апопласт и симплета, докато достигне ендодермата. По-нататъшното движение на Apoplast е невъзможно.
Различни видове структура на централния цилиндър на корена (първична структура): A-Diarch, B-Triirchny, е тетрархичен, полиархин. Видове A-B са характерни за дикотиятик, г-н в много едноетажни. 1 - Парцел на първична кора, 2 - първичен етаж, 3 - първичен xylem.
4 етапа на кореновия преход от първичната структура до вторична: 1) разграничават сермите на CAMBIA между първичните флола и ксилемите; 2) образуването на перикола на разлогането; 3) освобождаване от първичната кора; 4) Промяна на радиалното подреждане на обезпечението на проводимите тъкани.
Преходът от първичната структура на корена до вторичен 1-целевия етаж, 2-експлоатационен ксислам, 3 - cambier, 4 - перикола, 5 - endoderma, 6 - мезодерма, 7 - Rizoderma, 8 - Exoderma, 9-Real Xylem , 10 - вторичен плам, 11 - вторичен CORA, 12 - гълъб, 13 - FELLEM.
Схема на първична диференциация на проводящите тъкани в корена на ЧСИ 1 центродвигателен диференциат на XILEMA, 2 -epiderma, 3-лични рамки, 4-бендармени, 5-ин-линейни диференцирани елементи на ксиславе, 6-образни елементи на ксиславе, 7 -Проосревни диференцирани елементи на флола, 9-пик на меристем, 10-aknevial \u200b\u200bслучай.
Вторична промяна в корена Monocoa. Преобладаващото мнозинство от електронни растения основната структура на корена се задържа до края на живота. В същото време са назначени много елементи на корена. Дървен монооцентност (палмови дървета, дражети, yukki) в коренната корка на паренхимните клетки или от пейчицилетите възникват слой от меристема. Той образува редове от затворени проводими греди. След това в периферната част на паренхимата се появява нов слой от образователна тъкан на паренхимата на първичната кора. Този слой на меристема дава началото на нов ред проводящи греди. Така се случва удебеляване на корена.
Корените под налягане възникват върху различни органи на растенията - на стъбла, листа и корени. Очевидните корени, възникнали върху стъблото, се наричат \u200b\u200bStelerrodukuks, които са възникнали в корена - корен. Страничните и очевидните корени имат ендогенен произход, т.е. те са положени във вътрешните тъкани.
Заключването на страничния корен започва с разделянето на перило-клетки. В същото време на повърхността на Stela се образува месистематично туберкулово. След редица разделения на клетките на meristematic tuberca се случва страничен корен. Той има свой апик и случай. Потомството на страничния корен расте, прекъсва основната кора на корена на майката и се простира навън. Обикновено се появяват странични корени срещу елементите на ксилемите. Затова те са разположени правилно надлъжни редове по корена. Те възникват в абсорбционната зона или малко по-висока. Заключването на страничните корени се появява както акроптално, т.е. от основата на корена до върха.
Корените за налягане обикновено са поставени в тъкани, способни на по-меристематична активност: в Pericycle, Cambia, Mellogene. Ендогенното образуване на странични (и пресоване) корени има адаптивна стойност. Ако клонът се появи в върха, напредъкът на корена в почвата беше труден.
Дихотомното разклоняване в кореновата система на Пладен маланоид (Lycopodium clavatum) 1 - изотомиран дихотомно разклоняване на най-тънките корени
Mycorrhuses: А - Етототрофичен дъб Микориза, б, б - ендотрофан Микариза Ятришник. Nujna на корените на лупина
Коренни функции.Коренът е основният орган на най-високата инсталация. Функциите на корените са следните:
Почвите се абсорбират от почвата и минералните соли, разтворени в нея, транспортират ги стъпки, листа и репродуктивни органи. Функцията за всмукване се извършва чрез коренни косми (или микороиди), разположени в смукателната зона.
Благодарение на голяма сила, растението е фиксирано в почвата.
- При взаимодействието на водата йони на минерални соли и фотосинтезни продукти синтезират продуктите от първичен и вторичен метаболизъм.
- Под действието на кореновата налягане и прозрачни йони водни решения Минералите и органичните вещества съгласно корена на плавателните съдове KSILI се движат по нарастващия ток в стъблото и листата.
- Хранителните вещества (нишесте, инулин и др.) Са депозирани в основата.
- Корените се извършват чрез биосинтеза на вторични метаболити (алкалоиди, хормони и други бав).
- Синтезирани в мерматичните зони на корените на корените (Gibberllinins и т.н.) са необходими за растежа и развитието на надземните части на растението.
- Благодарение на корените, симбиозата се извършва с почвени микроорганизми - бактерии и гъби.
- Използването на корените има вегетативно възпроизвеждане на много растения.
10. Някои корени изпълняват функцията на респираторното тяло (чудовище, филодендрон и др.).
11. Корените на реда на растенията изпълняват функцията на "ранени" корените (Ficus banyan, Pandanus и др.).
12. Коренът е способен на метаморфоза (удебеляване на главния корен "корени" в моркови, магданоз и др.; Удебеляване на странични или притискащи корени форми на корен грудки в Далия, земни ядки, почистване и др., Съкращаване на корените в луковици).
Корен - аксиален орган, обикновено цилиндрична форма, с радиална симетрия с геотропизъм. Расте, докато топката на топлина се запази, покрита с основен случай. В корена, за разлика от бягството, листата никога не се образуват, но като бягство, кореновите клони, образувайки коренова система.
Коренната система е съвкупност от корените на едно растение. Естеството на кореновата система зависи от съотношението на растежа на основните, странични и очевидни корени.
^ Видове корени и коренови системи.При нудката на семето всички органи на растението са в детството. Началник или първи коренът се развива от покрив.Главният корен се намира в центъра на цялата коренова система, стволът служи като продължение на корена и заедно те представляват ос от първата поръчка. Наречен е парцел между главния корен и стъблото коренова врата.Този преход от стъблото към корена е забележим при различна дебелина на стъблото и корен: стъблото е по-дебело от корена. Парцел от коренна кормин до първите зародишни листа - наречени котиледи садделтънското коляноили хипокотилен. Страничните корени на следващите поръчки се отклоняват от главния корен към страните. Такава коренна система се нарича сантнева, много двупартитни растения, може да се разклонява. Разклонена коренова система е разнообразие от коренова система. Страничната част на корена се характеризира с факта, че новите корени са положени на известно разстояние от върховете и се образуват ендогенни - във вътрешните тъкани от предходния ред на основния корен поради активността на перикола. Колкото по-странични корени от главния корен, по-големите съоръжения на растението, следователно има специални агротехнически техники, които подобряват способността на основния корен да образуват странично, например, прищипване или гмуркам сеглавния корен върху л / 3 от дължината му. След гмуркане, известно време главният корен престава да расте дълго, докато страничните корени растат интензивно.
В растенията на дисфотрол, главният корен, като правило, е запазен през целия си живот, един боклук зародишен корен бързо умира, главният корен не се развива и от основата на бягството се образува подадкорени, които също имат клонове на първата, втора и т.н. поръчки. Такава коренна система се нарича bashed.Корените под налягане, като страна, са положени ендогенни. Те могат да бъдат оформени на стъбла и листа. Способността на растенията за развитие на очевидните корени се използва широко в производството на култури в растително възпроизвеждане на растения (възпроизвеждане на стотици и резници за листа). Резници за въздушен удар определят IVI, топола, клен, касис черно и др.; Листови резници - виолетов узбамбар, или сатполия, някои видове бегонии. Подземни резници на модифицирани издънки (коренища) Умножават мнозина лечебни растения, например, лилия на лилията може, купувайки лекарствени и други. Някои растения образуват много видими корени, когато дъното на стъблото (картофи, зеле, царевица и др.), По този начин създават допълнителна храна.
При най-високите спорове (растения, сандъци, папрати), главният корен изобщо не се случва, те се образуват само очевидните корени от корена. В много дикарпутични тревни коренични растения, главният корен често се отклонява и преобладава системата на очевидните корени, които се отдалечават от коренища (болни, коприва, щраус и д-р.).
На дълбочината на проникване в почвата, първото място принадлежи на коренната система на пръчката: дълбочината на реколтата на проникването на корен, според една информация, достига 120 m! Въпреки това, кореновата система на урината, притежаваща основното повърхност на корените, допринася за създаването на тревата и предупреждава ерозията на почвата.
Общата дължина на корените в кореновата система е различна, някои корени достигат няколко десетки и дори стотици километри. Например, дължината на пшеницата на всички коренни косми достига 20 км, а зимният ръж има обща дължина на корените на първия, втория и третия полет от над 180 км, и с добавянето на корените на четвърти поръчки - 623 км. Въпреки факта, че коренът расте целия си живот, нейният растеж е ограничен до влиянието на корените на други растения.
Степента на развитие на кореновите системи върху различни почви в различни природни зони. Така че, в пясъчните пустини, където са дълбоко разположени подземните води, корените на някои растения отиват на дълбочина 40 м и повече (Zlak Selin, просопис около семейството на боб и др.). Plates-eememera полу-пустини имат повърхносткоренната система, адаптирана към бързото поглъщане на раните на влагата, е доста достатъчна за бързо преминаване на всички фази на растителната растителност. На глина, лошо газирани субзоли на горската зона на Тайга, кореновата система на растенията с 90% е концентрирана в повърхностния слой на почвата (10-15 см), растенията имат "корени" (европейски смърч). Например, Saksaul има корени в различно време Години използват влага различни хоризонти.
Много важен фактор при разпределението на кореновата система - влажност. Посоката на корените отива в страната на по-голямата влажност, но във вода и в претоварените корените на почвата са разклонени много по-слаби.
Степента на развитие на кореновите системи, дълбочината на проникването на корен и други пластмасови характеристики на корена зависи от външните условия и в същото време са наследствено фиксирани зад всяка растителна инсталация.
^ Зоната на младия корен.В младия корен разграничават: 1) зоната на разделяне, покрита с корен; 2) зоната на разтягащите клетки или зоната на растеж; 3) зона за всмукване или коренна зона; 4) Проводима зона.
^ Division Zone.е върхът на корена, външен покрит корен калъфзащита на горната или апикална, апарат. На допир, младият връх на корена е хлъзгав, дължащ се на секретираните клетки на слузта. С повишаването на корена в дължината на слузта намалява триенето на върха на корена на почвата. Според изразяването на академик v.l. Комаров, основен случай "root oneam", тя защитава разделителните клетки на меристемата от механични повреди и също така контролира положителен геотропизъмсамата корен, т.е. допринася за растежа на корена и прониква в дълбините на почвата. Коренният случай се състои от живи паренхимни клетки, в които присъстват нишестени зърна. В случая има зона за разделяне, или корен бърз конуспредставени от първична образователна тъкан (meristem). В резултат на активното разделение на елитната мярка на корена се образуват всички други корени и тъкани. Подразделната зона на млад корен е само 1 mm. От други зони изглежда жълто.
^ Zensile зона,или зона на растеж,дължината на няколко милиметра е външно прозрачна, състои се от почти неоткрити, но разтягащи клетки в надлъжната посока. Клетките се увеличават по размер, вакулите се появяват в тях. Клетките се характеризират с висока обиколка. В зоната на разтягане се наблюдава диференцирането на първичните проводими тъкани и постоянната корена започва да се образува.
Над разтегната зона се намира всмукателна зона.Дължината му е 5 - 20 мм. Зоната на засмукване е представена от корен косми - епидермални клетки са се увеличили. Използване на коренни косми, абсорбция на водни почви и соли решения. Многобройните кормилни косми, толкова по-голяма е всмукателната повърхност на корена. С 1 mm на повърхността на корена могат да бъдат разположени около 400 коренни косми. Коренните косми са краткотрайни, живеят 10 - 20 дни, след което те умират. Дължината на коренните косми в различни растения от 0.5 - 1.0 cm. Младите коренни косми са оформени над зоната на опън и умират над смукателната зона, така че зоната на коренните косми непрекъснато се движи като корен и растението се увеличава, то Възможно е да се абсорбира вода и да се разтвори в него. хранителни вещества от различни почвени хоризонти.
Над началото на смукателната зона започва зона за задържане или странична коренова зона.Водата, погълната от решенията на корена и солите, се транспортират с дървени съдове до надземните части на растението.
Между кореновите зони няма остри граници и има постепенно преход.
6. Коренна метаморфоза. Тяхната биологична значимост. Микориза.Повечето растения в една и съща коренова система са ясно разграничени растеж и всмукващкрай. Лисличността обикновено е по-мощна, бързо удължават и напредват в почвата. Зоната на разтягане е добре изразена, а апикалните меристеми работят енергично. Суспанките, възникнали в големи количества върху корените на растеж, се удължават бавно и техните апикални меристеми почти престават да работят. Изсмукващият край изглежда спира в почвата и интензивно "смущава го".
Дървесните растения разграничават дебели skelette. и семеенкорени, на които краткотраен root mety. Комплектът настоятелно призовава, че непрекъснато се заменят взаимно да включват растеж и смучещи окончания.
Ако корените изпълняват специални функции, структурата им се променя. Нарича се рязко, наследствена фиксирана модификация на органа, причинена от променящите се функции, се нарича метаморфоза. Модификациите на канала са много разнообразни.
Корените на много растения образуват симбиоза с Gifs of почвени гъбички, наречени микодизъм("Mibocornia"). Mikoriza се формира на смучещи корени в абсорбционната зона. Компонентът на гъби улеснява корените за получаване на вода и минерални елементи от почвата, често GIF файлове на гъби заместват коренните косми. От своя страна гъбата получава въглехидрати и други хранителни вещества от растенията. Има два основни вида микороза. Gifs. етототрофнамикорозата образува капак, обгръщайки корена отвън. Екстрахикорозата е широко разпространена в дървета и храсти. Ендотрофнаmycorroge се намира главно в тревни растения. Endomicuriza се намира вътре в корена, GIF файла се въвеждат в клетките на кравенето на паренхима. Микотрофното хранене е много широко разпространено. Някои растения, като орхидея, не могат да съществуват без симбиоза с гъби.
На корените на бобовите растения има специални формации - военноморскив които бактериите са вмъкнати от рода ризобий. Тези микроорганизми са в състояние да абсорбират атмосферния молекулен азот чрез превръщането му в съответното състояние. Някои от веществата, синтезирани в глупаците, се абсорбират от растенията, бактериите, от своя страна, използват вещества, които се корени. Тази симбиоза е от голямо значение за селско стопанство. Фасулките благодарение на допълнителен източник на азот са богати на протеини. Те дават ценни продукти за храна и фуражи и обогатяват почвата с азотни вещества.
Много широко разпространено боядисване Корени. Обикновено те се сгъстяват и силно паренхимминизират. Извършени са много удебелени явни корени коренни конуси, или cornklubnia.(Джоргин, някои орхидея). Много често, отколкото се появяват за растения с коренна система, образование, което се нарича корнефлода. Главният корен и долната част на стъблото участват в образуването на корена. Морковът почти всички корен се компилират от корена, коренът на ряпа образува само най-ниската част от корена на корена ( фиг. 4.12).
Корени културни растения Той е възникнал в резултат на дълъг подбор. В routfolds, отглеждането на паренхима е силно развито и механичните тъкани изчезнаха. Морковите, магданозът и други чадърен панерм са силно развити на платието; Полета, репички и други кръстоцветни - в ксилен. В Бекла, резервните вещества се отлагат в паренхим, формиран от дейностите на няколко добавяща слоя на Камбия ( фиг. 4.12.).
Образуват се много растения за насилие и коренище прибиране, или договор корените ( фиг. 4.13, 1.). Те могат да съкращават и да издърпат бягството в почвата на оптималната дълбочина по време на лятото суша или зима Морозов. Корените за вкореняване са удебелени бази с напречни бръчки.
Респираторно Корени, или пневматични телефони (фиг. 4.13, 2.) Те се оформят в някои тропически дървесни растения, живеещи в условия на липса на кислород (таксодий, или блато кипарис; мангрово растения, които живеят на блатисти брегове на океанските брегове). Пневмофорите растат вертикално нагоре и са изхвърлени по повърхността на почвата. Чрез системата на дупки в тези корени, свързани с Arenhima, въздухът влиза в подводни органи.
Някои растения за поддържане на бягства във въздуха се образуват допълнителни справка Корени. Те се отклоняват от хоризонталните клони на короната и достигат повърхността на почвата, интензивно разклонена, превръщайки се в стълбове, поддържащи короната на дървото ( стълбовеrainan корени) ( фиг. 4.15, 2). Пешеходци Корените се отклоняват от долните участъци на стъблото, давайки стабилност стабилност. Те се формират в растения от мангрови гъсталаци, растителни съобщества, развиващи се на наводнени по време на въдица на тропическите брегове на океаните ( фиг. 4.15, 3.), както и в царевица ( фиг. 4.15, 1.). Формиран Ficus Rubmicing. шарковкорени. За разлика от стълбовете и колебания, те не се оценяват по произход, а от странични корени.
Фиг. 4.15. ^ Поддържани корени : 1 - скитащи корени от царевица; 2 - стълбове на Банян; 3 - скитащи корени на ризофорлас ( и т.н.- зона на прилива; от - заключваща зона; i л.- повърхността на rostive дъното).
Концепция за стрелба. Морфологично несъгласие на бягството. Възли и интермодони. Най-добрите стреля от бягство. Строителство и дейностСтроителство. Escape се нарича стъбло с листата и бъбреците, разположени на него.
Раздели на стъблата, върху които се развиват листата, повикващи възли.
Раздели на стъблото между двете най-близки възли наречени междудържаща.
Ъгълът между листа и горното сношение наречен синусов лист.
В синуса на листа образуват упоритен бъбрек. Escape се състои от повтарящи се места - metamers..
Един метад включва интерщерци, възел, лист и stubby бъбрек. Escape се нарича комплекс, състоящ се от стъбло и листа. Първичното бягство е положено в пъпка, където е поразяване. Бъбрекът се състои от зародична стъбла - епикотил, апикална меристем и една или повече листа програми (твърди от листата). При поглъщане на семена, скелетът се удължава. От апикални меристеми развиват нови листни страни, от лист Примредиев развиват листа и в синуси от листа Образувана монвейка. Този алгоритъм за развитие по време на формирането на системата за снимане на растението може да повтаря многократно.
Формиран бягство отличава възлите - част от бягството, където листът е свързан с стъблото; Интезий - част от бягството между възлите, обикновено е част от стъблото; Синусите за листата са ъгъл между листа и възходящата част на стъблото.
Част от бягството са бъбреци. Това е преди всичко, най-големият бъбрек, който представлява увеличаването на конуса. В синуси от листа Семенните растения са оформени стрес или странични бъбреци. Ако те развиват един над друг (орлови нокти, орех, Робиния и т.н.), след това се нарича сериен. Ако бъбреците се развият в синусите на листата наблизо, за приятел (слива, зърнени култури и т.н.), тогава те се наричат \u200b\u200bобезпечение. Бъбреците могат да образуват ендогенно в областта на Interdozy. Тези бъбреци се наричат \u200b\u200bочевидни.
Дърветата и храстите на студен и умерен климат са оформени зимуващи или почиват бъбреците, които често се наричат \u200b\u200bоко. От тези бъбреци, новите издънки развиват следващата година. Външните листа в тези бъбреци обикновено се превръщат в бъбречни скали, които предпазват вътрешните части на бъбреците от повреда.
Зима или почивка бъбреците се образуват в многогодишни билки, върху тези органи, които не умират за зимата, т.е. върху коренища, в основата на стъбла и др. Тези бъбреци се наричат \u200b\u200bбъбрек за обновяване. От тях през пролетта развива надземни издънки.
Всички гореспоменати бъбреци се наричат \u200b\u200bвегетативни. Такива бъбреци се състоят от върха, промишлени комплекти, инхаракции на интерстициал, лист Примредиев, над кои бъбречни държави могат да развият и подиграват листата.
От бъбреците, които нямат бъбречен приоритет, прост или кафяв бягството. Разликата се развива от бъбреците с приоритет на бъбреците бягството.
В допълнение, семена растенията имат повече генеративни бъбреци. Това са цъфтящи бъбреци и бъбреци, което поражда надалените. Те се различават от вегетативния външен вид. В допълнение към върха, пълнените интерстициални и нелепи единици, такива бъбреци имат премия, които дават на произхода на цветните части или части от конусите. На бъбреците, което поражда съцветия, се образуват цветни свлачища.
И накрая, има така наречени смесени бъбреци, от които дизайнерски издънки с цветя.
Морфологична характеристика Юджинът предполага описание на структурата на възлите, близките, бъбреците. Не забравяйте да посочите вида на регистрацията. За повечето растения това е друго - един лист се намира в възела, но може да бъде против или взаимен. Определен тип светло положение образува листа мозайка, която позволява по най-добрия начин Използвайте място, за да осигурите равномерно осветление на листа.
С процеса на растеж и развитието на бягство е свързано и разделянето на листа в три категории: долни листа, средни листа, Отгоре или каране на листа. В морфологичното описание на листата средните листа обикновено описват, но пълното морфологично описание изисква отделно описание на всички категории листа, защото дори средни листа На една стреля има различия. Този феномен се нарича хетерофилрия или несъответствие.
Топ нарастващо бягство - ръст на избягванията по дължина поради изменение на конуса на повишаването, инвестициите и растежа в основата на листата на инфлет. В процеса на модификация нарастващият конус се увеличава по дължина, става сложен и променя формата му.
Бъд. Това е успешен бягство. Състои се от меристематична ос, завършваща нарастващия конус (първичен ствол), а листният Progrdiyev (Incarpasses), т.е. от серия от мета мета камери. Листата по-долу са покрити под нарастващия конус. Така е подредена вегетативен бъбрек. В вегетативния репродуктивен бъбрек нарастващият конус се превръща в резервално цвете или залата на съцветие. Репродуктивните (цветни) бъбреци се състоят само от елементарно цвете или съцветия и нямат адаптация на фотосинтетични листа.
13. Метаморфизирани издънки.
Появата на тях често се свързва с изпълнението на функциите на удължаване на свободните продукти, прехвърлящи неблагоприятните условия на годината, вегетативно възпроизвеждане.
Коренище - Това е дългосрочно подземно бягство с хоризонтален, възходящ или вертикален растеж, който извършва функциите на натрупване на резервни продукти, подновяване, вегетативно размножаване. Коремът има намалени листа под формата на люспи, бъбреци, очевидни корени. Резервните продукти се натрупват в стеблото. Възходът и разклонението се случва точно като обикновен бягство. Кореното се отличава от корена чрез наличието на листа и липсата на корен случай на върха. Коремът може да бъде дълъг и тънък (питие) или къс и дебел. Всяка година са оформени годишни издънки от горните и скучни бъбреци. Старите части на кората постепенно изчезват. Растения с хоризонтални дълги коренища, образуващи много надземни издънки, бързо заемат голяма площ и ако това са плевели (пиене), борбата срещу тях е доста трудна. Такива растения се използват за фиксиране на пясъците (colosnyak, aristide). В Luragovychy зърнените култури с дълги хоризонтални коренища се наричат \u200b\u200bризаем (дивеч, менталист) и с късо храст (Тимофевка, предпазител). Корените се намират главно в многогодишни тревисти растения, но понякога храсти (лагери) и храст (Lingonberry, боровинки).
TUBER - Това е удебелена част от бягството, съвместимост на резервните продукти. Клубените са надземни и подземни.
Над главата Това е удебеляване на главната (колраби) или страна (тропически орхидеи) избяга и носи нормални листа.
Подземен клубер - сгъстяване на хипокотилен (циклама) или краткотрайно подземно бягство - аларма (картофи). Листата на подземния клуб са намалени, в синусите има бъбреци, наречени очи.
Главна колона - Това е краткотрайно пълзящо бягство, което служи за разпространение (изземване на територията) и вегетативно възпроизвеждане. Има дълги междинни и зелени листа. В възлите се образуват очевидни корени и от горния бъбрек - съкратено бягство (гнездо), което след алармата продължава независимо съществуване. Увеличава симпозиалния симпозиален колона. Надрежици, загубиха функцията на фотосинтезата и изпълнява главно функцията на вегетативната репродукция, понякога се наричат \u200b\u200bмустаци (ягоди).
Крушка - Това е скъсен ствол (дарение), носещ многобройни, плътно прикрепени листа и очевидни корени. На върха на дохода има бъбрек. Много растения (лук, лале, хиацинт и т.н.) се образуват от този бъбрек се образува въздушно излизане и се образува нова крушка от страничния бъбрек. Външни скали в повечето случаи суха, филм и извършване на защитна функция, вътрешни - месести пълни с резервни продукти. На формата на крушките има сферични, яйцевидни, сплетени и др.
Ревност Външно, прилича на крушка, но всички везни са сухи, а резервните храни се отлагат в стеблата (шафран, гладиолус).
Spinys. Те имат различен произход - от бягство (ябълково дърво, круша, завой, глог, глобус, цитрус), лист (барбарис) или частите му: Rakhis (астрагал), хляб (бяла акация), плака (Compace). Шпиони са характерни за растенията от горещи сухи местообитания.
Мустак Те се формират от бягство (грозде), лист или части: rakhis и няколко листа (грах), плочи (ранг.), Акценти (sassaparil). Служи за прикрепване към подкрепата.
FileCladium. - Това са плоски стружки, разположени в синусите на намалени листа. На тях са оформени цветя. Има предимно сухи местообитания (Iglitsa, Phillantus) в растенията. Режещи устройства - модифицирани листа, характерни за насекомите. Проучвания (Rosyanka, Mukholovka). Те имат формата на СПИН, врагчета, мехурчета или затръшване и опаковани плочи. Малки насекоми, влизащи в тях, умират, се разтварят, използвайки ензими и се консумират от растенията като предимно допълнителен източник на минерални вещества.
Коренът е подземен аксиален елемент на растенията, който е най-важната част от тях, главната им вегетативна власт. Благодарение на корена, растението е фиксирано в почвата и се държи там през целия жизнен цикъл, а също така осигуряват с вода, минерали и хранителни вещества, съдържащи се в нея. Има различни видове и видове корени. Всеки от тях има свои собствени отличителни характеристики. В тази статия ще разгледаме съществуващите видове корени, видовете коренови системи. Също така ще се запознаем с техните характерни черти.
Какви са видовете корени?
Стандартният корен се характеризира с филаментна или тясно цилиндрична форма. В много растения, в допълнение към основния (основен) корен, се развиват и други видове корени. По-подробно обмислете какво представляват.
Главен корен
Тази растително тяло се развива от ембрионалния корен на семената. Главният корен е винаги сам (други видове корени на растенията обикновено присъстват в множествено число). Той остава в завода през целия жизнен цикъл.
За корена се характеризира положителна геотропия, която се дължи на земната атракция, тя се задълбочава на субстрата вертикално надолу.
Поставяне на корени
Пентиналите наричат \u200b\u200bвидовете корени на растенията, които се образуват на други органи. Тези тела могат да бъдат стъбла, листа, издънки и т.н. Например, зърнените култури имат така наречените първични корени, които са положени в стъблото за ембриони. Те се развиват в процеса на покълване на семена почти едновременно с главния корен.
Също така има листни придадни типове корени (произтичащи от вкореняване на листата), инсулт или възли (оформен от коренища, надземни или подземни инсулт) и т.н., мощни корени се образуват на долните възли, които се наричат \u200b\u200bвъздух (или. \\ T справка).
Външният вид на очевидните корени причинява способността на растението да възпроизвежда вегетативно възпроизвеждане.
Странични корени
Странични обаждания, които възникват като страничен клон. Може да се образува както на основните и очевидни корени. В допълнение, те могат да бъдат разклонени от странично, в резултат на които се образуват страничните корени с по-високи поръчки (първо, второ и трето).
Големите странични органи се характеризират с напречен геотропизъм, т.е. техният растеж настъпва в практически хоризонтално положение или под ъгъл към повърхността на почвата.
Какво се нарича коренова система?
Коренната система нарича всички видове и видове корени, налични в едно растение (т.е. тяхната съвкупност). В зависимост от съотношението на растежа на основните, страничните и натискащите се корени, неговият тип и природа се определят.
Видове коренови системи
Ако основният корен е развит много добре и забележим сред корените на друг вид, това означава, че растението има система за пръти. Тя е присъща на предимно заводи.
Коренната система от този тип се отличава с дълбока покълване в почвата. Например корените на някои билки могат да проникнат в дълбочина 10-12 метра (Osay, засяване на люцерна). Дълбочината на проникване на корените на дърветата в някои случаи може да достигне 20 m.
Ако видимите корени, които се развиват в големи количества, са значителни, а основната характеризира бавния растеж, се оформя кореновата система, която се нарича урина.

Такава система се характеризира като правило, някои от тревните. Въпреки факта, че корените на системата на урината проникват, не са толкова дълбоки, като пръчката, те по-добре увеличават частиците на почвата в непосредствена близост до тях. Много богати кръвни и коренище зърнени билки, образуващи богато количество тънки корени, са широко използвани за осигуряване на клисури, почви на склонове и т.н., огънят на най-добрите билки е огън, овесена каша и др.

Модифицирани корени
В допълнение към типичните, описани по-горе, има и други видове корени и коренови системи. Те се наричат \u200b\u200bмодифицирани.
Мигащи корени
Пейнтър включва корен и съкращение.
Коренният корен е удебеляване на главния корен поради отлагането на хранителни вещества в него. Също така, дъното на стъблото участва и в образуването на корена. Състои се повече от басейн от основна тъкан. Примери за корените могат да служат като магданоз, репички, моркови, цвекло и др.

Ако страничните и очевидни корени са удебелени корени, тогава те се наричат \u200b\u200bклубени от коренбал (конуси). Те са разработени в картофи, баня, Далия и др.
Въздушни корени
Това са странични корени, които растат в надземната част. В редица тропически растения. Вода и кислород абсорбират от въздушен. Има тропически растения, които растат в условия на липса на минерали.

Респираторни корени
Това разнообразие от странични корени, които растат, повдигащи се над повърхността на субстрата, вода. Такива видове корени са оформени в растения, растящи на твърде навлажнени почви, в условия на блатото. С помощта на такива корени, растителността получава липсващ кислород от въздуха.
Поддържащи (Sharkovid) корени
Тези видове дървета са характерни за големи породи (бук, елм, топола, тропически и т.н.) са триъгълни вертикални растения, образувани от странични корени и преминават на повърхността на почвата или над нея. Те също се наричат \u200b\u200bгладко, тъй като приличат на дъските, които се облягат към дървото.
Корен сукане (gautory)
Това е вида на допълнителните очевидни корени, които се развиват по стъблото на къщите растения. С тяхната помощ растенията имат способността да се прикрепят към специфична подкрепа и да се издигнат (разкъсано). Такива корени са на разположение, например във верига Ficus, бръшлян и др.

Интензивни (прецизни) корени
Характеристика за растенията, чийто корен е рязко намален в надлъжната посока в основата. Пример може да сервира растения с крушки. Интензивните корени осигуряват крушки и вкоренени вдлъбнатина в почвата. В допълнение, тяхното присъствие се причинява от плътна форма на гнездата (например при глухарче) до земята, както и подземното положение на вертикалния корен и кореновата шийка.
Mikoriza (mribocornia)
Микоридът се нарича симбиоза (взаимно изгодно съжителство) на корените на висшите растения с гъби хифи, които ги накисват, изпълнявайки функциите на коренни косми. Гъбите осигуряват растения с вода и хранителни вещества, разтворени в нея. Растенията, от своя страна, дават гънки, необходими за препитанията си органични вещества.
Присъщ на микорнизма към корените на много висши растения, особено дървесни.
Бактериални клубени
Това са модифицирани странични корени, които са адаптирани към симбиотично съжителство с фиксиране на азотни бактерии. Образуването на мускулите се дължи на проникването в младите корени. Такова взаимноизгодно съжителство позволява на растенията да получат азот, който бактериите се превеждат от въздуха към формата, която им е на разположение. Бактериите също са снабдени със специални местообитания, където могат да функционират, без да се конкурират с други видове бактерии. Освен това те използват вещества, присъстващи в корените на растителността.

Бактериалните клубени за растения от семейството на бобовите растения се характеризират, които се използват широко като амелиори в ротации на културите, за да обогатяват почвения азот. Най-добрите растения за фиксиране на азот са билките на род-зеления боб, като Lucerne Blue и жълто, червено и еспарц, рога и др.
В допълнение към горната метаморфоза, има и други видове корени, като корени на архивирането (помагат за укрепване на стеблото), пешеходни корени (помагат на растенията, които не потъват в течни мръсотия) и кореновите братя и сестри (имат нанасяне на бъбреци и да осигурят вегетативно възпроизвеждане) .









