Šaknų ir šaknų sistemos. Šoninių šaknų formavimas
Filogenetiniu požiūriu šaknis atsirado vėliau nei stiebas ir lapas - ryšium su augalų perėjimu į gyvenimą sausumoje ir tikriausiai atsirado iš šaknis primenančių požeminių šakų. Šaknis neturi lapų ar pumpurų, išdėstytų tam tikra tvarka. Jam būdingas viršūninis ilgio augimas, jo šoniniai padariniai kyla iš vidinių audinių, augimo taškas yra padengtas šaknies dangteliu. Šaknų sistema formuojasi visą augalo organizmo gyvenimą. Kartais šaknis gali tarnauti kaip maistinių medžiagų tiekimo vieta. Šiuo atveju jis yra modifikuotas.
Šaknų tipai
Pagrindinė šaknis susidaro iš embriono šaknies, kai sėkla sudygsta. Iš jo tęsiasi šoninės šaknys.
Ant stiebų ir lapų vystosi atsitiktinės šaknys.
Šoninės šaknys yra bet kurios šaknies šakos.
Kiekviena šaknis (pagrindinė, šoninė, atsitiktinė) turi galimybę išsišakoti, o tai žymiai padidina šaknų sistemos paviršių, o tai prisideda prie geresnio augalo stiprinimo dirvožemyje ir jo mitybos gerinimo.
Šaknų sistemų tipai
Yra du pagrindiniai šaknų sistemų tipai: pagrindinė, su gerai išvystyta pagrindine šaknimi ir pluoštinė. Pluoštinė šaknų sistema susideda iš daugybės to paties dydžio atsitiktinių šaknų. Visa šaknų masė susideda iš šoninių ar atsitiktinių šaknų ir atrodo kaip skiltelė.
Labai šakota šaknų sistema sudaro didžiulį sugeriantį paviršių. Pavyzdžiui,
- bendras žieminių rugių šaknų ilgis siekia 600 km;
- šaknų plaukų ilgis - 10 000 km;
- bendras šaknies paviršius - 200 m 2.
Tai daug kartų viršija antžeminės masės plotą.

Jei augalas turi gerai apibrėžtą pagrindinę šaknį ir vystosi atsitiktinės šaknys, tada susidaro mišri šaknų sistema (kopūstai, pomidorai).
Išorinė šaknies struktūra. Vidinė šaknies struktūra
Šaknų zonos

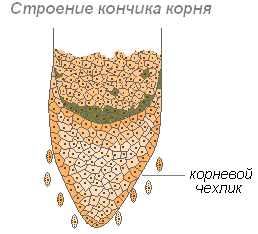
Šaknies dangtelis
Šaknis ilgėja viršūnėje, kur yra jaunos ugdomojo audinio ląstelės. Auganti dalis yra padengta šaknies dangteliu, kuris apsaugo šaknies galiuką nuo pažeidimų ir palengvina šaknies judėjimą per dirvą augimo metu. Pastaroji funkcija atliekama dėl to, kad šaknies dangtelio išorinės sienos turi būti padengtos gleivėmis, o tai sumažina trintį tarp šaknies ir dirvožemio dalelių. Jie netgi gali atstumti dirvožemio daleles. Šaknies dangtelio ląstelės yra gyvos ir dažnai turi krakmolo grūdelių. Dangtelio ląstelės nuolat atsinaujina dėl dalijimosi. Dalyvauja teigiamose geotropinėse reakcijose (šaknų augimo kryptis Žemės centro link).
Dalijimosi zonos ląstelės aktyviai dalijasi; šios zonos ilgis skirtingoms rūšims ir to paties augalo šaknims nėra vienodas.
Tempimo zona (augimo zona) yra už padalijimo zonos. Šios zonos ilgis neviršija kelių milimetrų.
Kai linijinis augimas baigiamas, prasideda trečiasis šaknų formavimosi etapas - jo diferenciacija, susidaro ląstelių diferenciacijos ir specializacijos zona (arba šaknų plaukelių ir absorbcijos zona). Šioje zonoje jau išskiriamas išorinis epiblemos sluoksnis (šakniastiebiai) su šaknies plaukeliais, pirminės žievės sluoksnis ir centrinis cilindras.
Šaknies plaukų struktūra
Šaknies plaukai yra labai pailgi išorinių ląstelių, apimančių šaknį, ataugos. Šaknies plaukų skaičius yra labai didelis (1 mm 2 nuo 200 iki 300 plaukų). Jų ilgis siekia 10 mm. Plaukai formuojasi labai greitai (jaunuose obelų daigais per 30-40 valandų). Šaknies plaukai yra trumpalaikiai. Jie miršta po 10-20 dienų, o jauni auga šaknies dalyje. Tai užtikrina šaknų naujų dirvožemio horizontų vystymąsi. Šaknis nuolat auga, formuoja vis daugiau šaknų plaukų sričių. Plaukai gali ne tik sugerti paruoštus medžiagų tirpalus, bet ir padėti ištirpinti kai kurias dirvožemio medžiagas, o tada jas įsiurbti. Šaknies sritis, kurioje šaknų plaukai išnyko, kurį laiką sugeba sugerti vandenį, tačiau vėliau ji uždengiama kamščiu ir praranda šį gebėjimą.
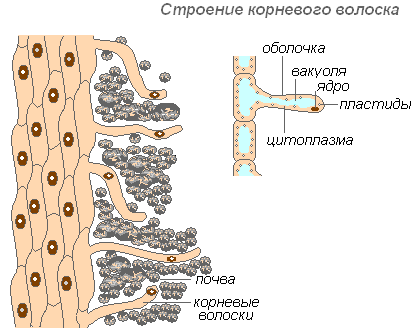
Plaukų apvalkalas yra labai plonas, todėl lengviau įsisavinamos maistinės medžiagos. Beveik visą plaukų ląstelę užima vakuolė, apsupta plono citoplazmos sluoksnio. Branduolys yra ląstelės viršuje. Aplink ląstelę susidaro gleivinis apvalkalas, kuris skatina šaknų plaukelių sukibimą su dirvožemio dalelėmis, o tai pagerina jų kontaktą ir padidina sistemos hidrofiliškumą. Absorbciją skatina šaknų plaukuose išsiskiriančios rūgštys (anglies, obuolių, citrinos), kurios tirpdo mineralines druskas.
Šaknies plaukai taip pat atlieka mechaninį vaidmenį - jie tarnauja kaip šaknies viršūnės atrama, kuri praeina tarp dirvožemio dalelių.
Po mikroskopu, skersinėje šaknies atkarpoje absorbcijos zonoje matoma jos struktūra ląstelių ir audinių lygiu. Šaknies paviršiuje yra šakniastiebis, žemiau - žievė. Išorinis žievės sluoksnis yra eksoderma, į vidų - pagrindinė parenchima. Jo plonasienės gyvos ląstelės atlieka saugojimo funkciją, atlieka maistinių medžiagų tirpalus radialine kryptimi - nuo siurbiamojo audinio iki medienos indų. Jie taip pat sintezuoja daugybę organinių medžiagų, gyvybiškai svarbių augalui. Vidinis žievės sluoksnis yra endoderma. Maistiniai tirpalai iš žievės į centrinį cilindrą per endodermines ląsteles praeina tik per ląstelių protoplastą.

Žievė supa centrinį šaknies cilindrą. Jis ribojasi su ląstelių sluoksniu, kuris ilgą laiką išlaiko savo gebėjimą dalintis. Tai yra dviratis. Dviračio ląstelės sukelia šonines šaknis, atsitiktinius pumpurus ir vidurinio ugdymo audinius. Į vidų nuo dviračio, šaknies centre, yra laidūs audiniai: masto ir medžio. Kartu jie sudaro radialiai laidų ryšulį.
Šaknies laidumo sistema praleidžia vandenį ir mineralus nuo šaknies iki stiebo (srovė aukštyn), o organinės medžiagos - nuo stiebo iki šaknies (žemyn srovė). Jį sudaro kraujagyslių pluoštiniai ryšuliai. Pagrindinės ryšulio sudedamosios dalys yra floemo sekcijos (išilgai kurių medžiagos juda į šaknį) ir ksilemas (išilgai kurių medžiagos juda iš šaknies). Pagrindiniai floemo laidieji elementai yra sietiniai vamzdeliai, ksilemos - trachėja (indai) ir tracheidos.
Šaknies gyvybiniai procesai
Šakninis vandens transportas
Vandens įsisavinimas šaknų plaukeliais iš dirvožemio maistinių medžiagų tirpalo ir nešimas radialine kryptimi išilgai pirminės žievės ląstelių per endodermos praėjimo ląsteles iki radialiai laidžiojo ryšulio ksilemos. Vandens sugerties šaknies plaukeliais intensyvumas vadinamas įsiurbimo jėga (S), jis lygus osmosinio (P) ir turgoro (T) slėgio skirtumui: S = P-T.
Kai osmosinis slėgis yra lygus turgoro slėgiui (P = T), tada S = 0, vanduo nustoja tekėti į šaknies plauko ląstelę. Jei dirvožemio maistinių medžiagų tirpale medžiagų koncentracija yra didesnė nei ląstelės viduje, tada vanduo paliks ląsteles ir įvyks plazmolizė - augalai nudžius. Šis reiškinys pastebimas esant sausam dirvožemiui, taip pat per daug tręšiant mineralinėmis trąšomis. Šaknies ląstelių viduje šaknies čiulpimo jėga didėja nuo šakniastiebio link centrinio cilindro, todėl vanduo juda išilgai koncentracijos gradiento (ty iš didesnės koncentracijos vietos į mažesnės koncentracijos vietą) ir sukuria šaknies spaudimą , kuris pakelia vandens stulpelį išilgai ksilemo indų, sudarant aukštyn srovę. Tai galima rasti ant lapų neturinčių pavasario kamienų, kai nuimama „sula“, arba ant nupjautų medžių kelmų. Vandens nutekėjimas iš medžio, šviežių kelmų, lapų vadinamas augalų „verkimu“. Žydėdami lapai taip pat sukuria čiulpimo jėgą ir pritraukia vandenį prie savęs - kiekviename inde susidaro ištisinė vandens stulpelis - kapiliarų įtempimas. Šaknies slėgis yra apatinis vandens srovės variklis, o lapų čiulpimo jėga - viršutinė. Tai galima patvirtinti atliekant paprastus eksperimentus.
Vandens absorbcija per šaknis
Tikslas: išsiaiškinti pagrindinę šaknies funkciją.
Ką mes darome: augalas, užaugintas ant šlapių pjuvenų, nupurtykite šaknų sistemą ir įdėkite šaknis į stiklinę vandens. Supilkite ploną augalinio aliejaus sluoksnį ant vandens, kad apsaugotumėte jį nuo garavimo ir pažymėkite lygį.

Ką mes stebime: per dieną ar dvi vanduo indelyje nukrito žemiau ženklo.

Rezultatas: todėl šaknys įsiurbė vandenį ir pakėlė į lapus.
Galima atlikti dar vieną eksperimentą, įrodantį, kad šaknis įsisavina maistines medžiagas.
Ką mes darome: Nupjaukite augalo stiebą, palikdami 2–3 cm aukščio kelmą. Ant kelmo uždėkite 3 cm ilgio guminį vamzdelį ir ant viršutinio galo uždėkite 20–25 cm aukščio išlenktą stiklinį vamzdelį.
Ką mes stebime: vanduo stikliniame vamzdyje kyla ir teka.
Rezultatas: tai įrodo, kad šaknis sugeria vandenį iš dirvos į stiebą.

Ar vandens temperatūra turi įtakos šaknies vandens absorbcijai?
Tikslas: sužinokite, kaip temperatūra veikia šaknies darbą.
Ką mes darome: viena stiklinė turi būti su šiltu vandeniu (+ 17-18 ° C), o kita-šaltu (+ 1-2 ° C).
Ką mes stebime: pirmuoju atveju vanduo išleidžiamas gausiai, antruoju - mažai arba visiškai sustoja.
Rezultatas: tai įrodymas, kad temperatūra daro didelę įtaką šaknies veikimui.

Šiltas vanduo aktyviai įsigeria į šaknis. Šaknų slėgis pakyla.

Šaltas vanduo prastai įsigeria į šaknis. Šiuo atveju šaknų slėgis sumažėja.
Mineralinė mityba
Fiziologinis mineralų vaidmuo yra labai svarbus. Jie yra organinių junginių sintezės pagrindas, taip pat veiksniai, keičiantys koloidų fizinę būseną, t.y. tiesiogiai veikia protoplasto metabolizmą ir struktūrą; tarnauja kaip biocheminių reakcijų katalizatoriai; paveikti ląstelių turgoro ir protoplazmos pralaidumą; yra augalų organizmų elektrinių ir radioaktyviųjų reiškinių centrai.
Nustatyta, kad normalus augalų vystymasis įmanomas tik tuo atveju, jei maistiniame tirpale yra trys nemetalai - azotas, fosforas ir siera ir - ir keturi metalai - kalis, magnis, kalcis ir geležis. Kiekvienas iš šių elementų turi individualią reikšmę ir negali būti pakeistas kitu. Tai makroelementai, jų koncentracija augale yra 10 -2–10%. Normaliam augalų vystymuisi reikalingi mikroelementai, kurių koncentracija ląstelėje yra 10 -5 -10 -3%. Tai yra boras, kobaltas, varis, cinkas, manganas, molibdenas ir tt Visi šie elementai yra dirvožemyje, tačiau kartais jų nepakanka. Todėl į dirvą įterpiamos mineralinės ir organinės trąšos.
Augalas auga ir vystosi normaliai, jei visos reikalingos maistinės medžiagos yra šaknis supančioje aplinkoje. Dirvožemis yra tokia terpė daugumai augalų.
Kvėpuojančios šaknys
Normaliam augalo augimui ir vystymuisi tai būtina Grynas oras... Patikrinkime, ar taip yra?
Tikslas: ar šaknims reikia oro?
Ką mes darome: paimkite du identiškus indus su vandeniu. Į kiekvieną indą įdėsime besivystančius sodinukus. Mes kiekvieną dieną prisotiname vandens viename iš indų, naudodami purškimo buteliuką. Ant antrojo indo vandens paviršiaus užpilkite ploną augalinio aliejaus sluoksnį, nes tai lėtina oro srautą į vandenį.

Ką mes stebime: po kurio laiko augalas antrame inde nustoja augti, nudžiūsta ir galiausiai žūsta.

Rezultatas: augalo mirtis įvyksta dėl oro trūkumo, reikalingo šaknies kvėpavimui.
Šaknų modifikacijos
Kai kurie augalai kaupia atsargines maistines medžiagas šaknyse. Juose kaupiasi angliavandeniai, mineralinės druskos, vitaminai ir kitos medžiagos. Tokios šaknys stipriai auga ir įgyja neįprastą išvaizdą. Šaknis ir stiebas dalyvauja formuojant šakniavaisius.
Šaknys
Jei kaupiamosios medžiagos kaupiasi pagrindinėje šaknyje ir pagrindinio ūglio stiebo apačioje, susidaro šakniavaisiai (morkos). Šaknis formuojantys augalai dažniausiai yra bienalės. Pirmaisiais gyvenimo metais jie nežydi ir šakniavaisiuose kaupia daug maistinių medžiagų. Antra, jie greitai žydi, naudodamiesi sukauptomis maistinėmis medžiagomis, ir sudaro vaisius bei sėklas.
Šakniagumbiai

Jurliuose atsarginės medžiagos kaupiasi atsitiktinėse šaknyse, sudarydamos šakniagumbius.
Bakteriniai mazgeliai
Savotiškai pasikeitė dobilų, lubinų ir liucernos šoninės šaknys. Bakterijos nusėda jaunose šoninėse šaknyse, o tai palengvina dujinio azoto įsisavinimą dirvožemio ore. Tokios šaknys įgauna mazgelius. Šių bakterijų dėka šie augalai gali gyventi azoto neturtingose dirvose ir padaryti juos derlingesnius.
Sustingęs

Netoli potvynio rampos išsivysto pakrikusios šaknys. Jie laiko didelius lapuotus ūglius aukštai virš vandens ant netvirtos purvinos žemės.
Oras

Atogrąžų augalai, gyvenantys ant medžių šakų, vysto oro šaknis. Jie dažnai randami orchidėjose, bromeliadose ir kai kuriuose paparčiuose. Oro šaknys laisvai kabo ore, nepasiekia žemės ir sugeria drėgmę, kuri patenka ant jų nuo lietaus ar rasos.
Atsitraukiantis

Svogūninėse ir gumbasvogūniuose, pavyzdžiui, krokuose, tarp daugybės gijinių šaknų yra keletas storesnių, vadinamųjų atsitraukiančių, šaknų. Susitraukusios tokios šaknys gumbasvogūnius traukia giliau į dirvą.
Stulpelis

Fikusas vysto stulpines oro šaknis arba atramines šaknis.
Dirvožemis kaip šaknų buveinė
Dirvožemis augalams yra terpė, iš kurios jis gauna vandens ir maistinių medžiagų. Mineralinių medžiagų kiekis dirvožemyje priklauso nuo specifinių pirminių uolienų savybių, organizmų aktyvumo, nuo pačių augalų gyvenimo, nuo dirvožemio tipo.
Dirvožemio dalelės konkuruoja su šaknimis dėl drėgmės, išlaikydamos ją savo paviršiuje. Tai vadinamasis surištas vanduo, kuris yra suskirstytas į higroskopinį ir plėvelinį vandenį. Jį laiko molekulinės traukos jėgos. Augalui prieinamą drėgmę vaizduoja kapiliarinis vanduo, susikaupęs mažose dirvos porose.
Tarp dirvožemio drėgmės ir oro fazės atsiranda antagonistiniai santykiai. Kuo daugiau didelių porų dirvožemyje, tuo geresnis šių dirvožemių dujų režimas, tuo mažiau drėgmės dirvožemis sulaiko. Palankiausias vandens ir oro režimas palaikomas struktūriniuose dirvožemiuose, kur vanduo ir oras yra vienu metu ir netrukdo vienas kitam - vanduo užpildo kapiliarus struktūrinių agregatų viduje, o oras užpildo dideles poras tarp jų.
Augalo ir dirvožemio sąveikos pobūdis iš esmės yra susijęs su dirvožemio sugerties gebėjimu - gebėjimu išlaikyti ar surišti cheminius junginius.
Dirvožemio mikroflora suskaido organines medžiagas į paprastesnius junginius, dalyvauja formuojant dirvožemio struktūrą. Šių procesų pobūdis priklauso nuo dirvožemio tipo, cheminė sudėtis augalų liekanas, fiziologines mikroorganizmų savybes ir kitus veiksnius. Dirvožemio struktūros formavime dalyvauja dirvožemio gyvūnai: anelidės, vabzdžių lervos ir kt.
Derinant biologinius ir cheminius procesus dirvožemyje, susidaro sudėtingas organinių medžiagų kompleksas, kurį vienija terminas „humusas“.
Vandens kultūros metodas
Kokios druskos augalui reikalingos ir kokį poveikį jos turi augimui ir vystymuisi, buvo nustatyta eksperimentuojant su vandens augalais. Vandens kultūros metodas yra augalų auginimas ne dirvožemyje, bet mineralinių druskų vandeniniame tirpale. Priklausomai nuo eksperimento tikslo, galite pašalinti atskirą druską iš tirpalo, sumažinti arba padidinti jo kiekį. Nustatyta, kad azoto turinčios trąšos skatina augalų, kuriuose yra fosforo, augimą - ankstyvą vaisių nokinimą, o kalio turinčios - greičiausią organinių medžiagų nutekėjimą iš lapų į šaknis. Šiuo atžvilgiu trąšas, kuriose yra azoto, rekomenduojama naudoti prieš sėją arba pirmoje vasaros pusėje, kuriose yra fosforo ir kalio - antroje vasaros pusėje.
Naudojant vandens kultūrų metodą, buvo galima nustatyti ne tik augalo makroelementų poreikį, bet ir išsiaiškinti įvairių mikroelementų vaidmenį.
Šiuo metu pasitaiko atvejų, kai augalai auginami hidroponikos ir aeroponikos metodais.
Hidroponika - augalų auginimas konteineriuose, pripildytuose žvyro. Maistinių medžiagų tirpalas, kuriame būtinus elementus, tiekiamas į indus iš apačios.
Aeroponika yra oro augalų kultūra. Taikant šį metodą, šaknų sistema yra ore ir automatiškai (kelis kartus per valandą) purškiama silpnu maistinių druskų tirpalu.
ORGANOGRAFIJA
Organografija - gyvo organizmo organų aprašymas.
Vargonas- organizmo dalis, diferencijuota morfologiškai ir funkciškai. Atsižvelgiant į dvi pagrindines funkcijas: tam tikro augalo gyvybės palaikymą ir dauginimąsi tolesnėse kartose, išskiriami vegetaciniai ir generaciniai (reprodukciniai) organai.
Pirma, mes ištirsime vegetatyvinius organus, kurie sudaro ūglių sistemą ir šaknų sistemą. Šaknies sistemos elementas yra šaknis.
Šaknis
Šaknis evoliuciškai atsirado vėliau nei ūgliai ir kilo iš pirmųjų sausumos augalų - rinofitų - rizoidų.
Skirtingos šaknies dalys atlieka skirtingas funkcijas ir joms būdingos tam tikros morfologinės ypatybės. Šios dalys vadinamos zonomis. Jaunos šaknų zonos: padalijimo zona, augimo ar išplėtimo zona, absorbcijos arba siurbimo zona ir laidumo zona.
Išorinė viršūninė meristema yra padengta šaknies dangteliu. Gyvos dangtelio parenchiminės ląstelės gamina gleives. Gleivinė medžiaga yra labai hidratuotas pektino pobūdžio polisacharidas. Jis kaupiasi diktozomų pūslelėse, kurios, susiliejusios su plazmolemma, išleidžia jį į tarpą tarp plazmolemmos ir ląstelės membranos. Tada gleivės išeina į ląstelės membranos paviršių, kur susidaro maži lašeliai.
Dėl šios gleivėtos dangos dirvožemio dalelės prilimpa prie šaknies galiuko ir šaknų plaukų. Gleivės tarnauja kaip absorbuojantis paviršius jonų mainams. Gleivinė apsaugo šaknies galiuką nuo kenksmingų medžiagų dirvožemyje ir nuo išdžiūvimo sukuria palankias sąlygas naudingoms bakterijoms įsikurti.
Kai šaknis auga ir šaknies dangtelis yra stumiamas į priekį, ląstelės, esančios dangtelio periferijoje, nusileidžia. Ląstelių gyvenimo trukmė nuo atsiradimo iki pleiskanojimo yra 4–9 dienos, priklausomai nuo dangtelio ilgio ir augalo tipo.
Šaknies dangtelis valdo šaknies georecepciją. Gravitacijos suvokimo vieta dangtelyje yra centrinė ląstelių kolona - columella, kurioje amiloplastai veikia kaip statolitai (gravitacijos receptoriai).
Viršūninę meristemą sudaro mažos (10 - 20 µm skersmens) daugialypės ląstelės, turinčios tankią citoplazmą ir didelius branduolius.
Sporiniai augalai turi mono-pradinį šaknies viršūnės tipą.
Sėklinių augalų šaknyse aprašomi du pagrindiniai viršūninės organizacijos tipai. Vienoje iš jų visos sritys turi bendrus inicialus. Tai atviras viršūninių meristemų tipas, kuris tikriausiai yra filogenetiškai primityvus. Antrojo tipo atveju kiekviena iš trijų dalių, t.y. centrinis (arba laidus) cilindras, pirminė žievė ir šaknies dangtelis turi savo inicialus. Tai uždaras viršūninių meristemų tipas.
Inicialai, atsakingi už ląstelių augimo organizavimą pirmaisiais etapais, paprastai praranda mitozinį aktyvumą vėlesniuose šaknų augimo etapuose. Vietoj to, ląstelės, esančios šiek tiek giliau šaknies kūne, demonstruoja mitozinį aktyvumą. Šie stebėjimai paskatino formuoti poilsio centro teoriją viršūninėje meristemoje. Remiantis šia teorija, labiausiai nutolusios ląstelės (tolimiausia organo dalis, esanti toliausiai nuo centrinio taško: apatinis šaknies galas) šaknies kūne (pirmieji pleromos ir peribleles inicialai) dalijasi retai, turi maždaug tą patį dydis ir žemas NK ir baltymų sintezės greitis. Poilsio centras yra pusrutulio arba disko formos, skiriasi pagal tūrį, priklausomai nuo šaknies dydžio. Miegančios centro ląstelės dalijasi apie 10 kartų lėčiau nei kaimyninės ląstelės.
Šaknyse, eksperimentiškai pažeistas, poilsio centras sugeba atkurti meristemą. Tai yra, ramybės centras vaidina esminį vaidmenį formuojant struktūrą ir vystant šaknį.
Šaknies galiuko bruožas yra išilginių ląstelių eilučių, kilusių iš viršūninės meristemos, išdėstymas.
Šakninės viršūninės meristemos sluoksniai:
Dermatogenas(Gr. Derma - oda, genos - gimimas) - viršutinis viršūninės meristemos sluoksnis, diferencijuojasi į rizodermą. Tinkamos formos dermatogeninės ląstelės.
Pleroma(Graikų pleroma - įdaras) - centrinė viršūninės meristemos dalis, diferencijuojasi į centrinio cilindro ląsteles. Ląstelės yra pailgos ašiai.
Periblema(Graikų periblema - viršelis) - viršūninės meristemos sluoksnis tarp dermatogeno ir pleromos, diferencijuojasi į pirminės žievės ląsteles.
Pirminė žievė diferencijuojasi tempimo zonos lygyje. Silpnėjimo zonoje pirminė pluta sudaro didžiąją skerspjūvio dalį. Jis vadinamas „pirminiu“ dėl kilmės iš pirminės meristemos, „pluta“ - dėl periferinės vietos.
Išorinės pirminės žievės ląstelės, esančios po rizodermu, vadinamos eksoderma. Vidinis pirminės žievės sluoksnis yra endoderma.
Didžioji dalis pirminės žievės yra mezoderma. Mezodermą sudaro parenchiminės ląstelės, esančios laisvai, o tai skatina šaknų ląstelių vėdinimą.
Gimnašakių ir daugumos dviskilčių šaknys anksti numeta pirminę žievę. Jų pirminės žievės ląstelės išlieka parenchiminės. Priešingai, pirminė vienaląsčių žievė išlieka visą šaknies gyvenimą, o jos ląstelės sudaro antrines lignifikuotas membranas.
Šaknies centrą užima ašinis cilindras - stela. Pradeda diferencijuoti augimo zonoje. Ašinio cilindro formavimas prasideda nuo jo išorinio sluoksnio susidarymo - vienam dviračiui. Periciklas priklauso pirminiams šoniniams meristemams, jo ląstelės ilgą laiką išlaiko meristematinį aktyvumą. Paprastai dviratį vaizduoja vienas ląstelių sluoksnis, nors jis gali būti daugiasluoksnis.
Daugumoje sėklinių augalų šoninės šaknys dedamos į dviratį ir atsiranda atsitiktinių pumpurų užuomazgos. Rūšims, kurių augimas yra antrinis, dviratis dalyvauja formuojant kambiumą ir sukuria pirmąjį hellogeno sluoksnį.
Po dviračiu dedamos prokambi ląstelės, iš kurių susidaro pirminė floema, o vėliau ir pirminė ksilema. Ksilemo spindulių skaičius įvairiose rūšyse svyruoja nuo dviejų, trijų, keturių iki kelių. Pagal ksilemo spindulių skaičių šaknys vadinamos diarchu, triarchu, tetrarchu, polyarchu. Tetrarcho šaknys laikomos pagrindinėmis. Vienaląsčiai turi poliarchinę stelą.
Taigi, bendras planas pirminė šaknies struktūra ir audinių morfologinė diferenciacija rodo aukštą šaknies specializaciją ir prisitaikymą prie vandens ir mineralų absorbcijos ir transportavimo funkcijų.
Šoninių šaknų formavimas
Šoninės šaknys atsiranda centrinio cilindro periferijoje skirtingais atstumais nuo viršūninės meristemos. Kadangi jie dedami į giliai esančius šaknies audinius, jų kilmė vadinama endogenine (iš graikų kalbos „gimė viduje“). Šoninės gimnastikos ir angiospermos šaknys dedamos į dviratį. Periciklinės ląstelės, dalyvaujančios šoninių šaknų formavime, yra ragenos lankas.
Endoderma taip pat gali sudaryti kelis šaknies pradų ląstelių sluoksnius.
Diarchinėje šaknyje šoninė šaknis yra tarp floemos ir ksilemos, triarchinėje ir tetrarchinėje - priešais ksilemą, poliarchinė vienaląsčių šaknis - priešais floemą. Dviračio ląstelės dalijasi į skirtingomis kryptimis... Dėl to susidaro mažas šaknies gumbas. Padidėjęs dydis, šaknies pumpuras spaudžia endodermą. Tačiau endodermos lukštai yra kieti. Todėl endoderma išsipučia. Endodermio išsipūtimas vadinamas šaknies kišene.
Šaknies pradžia praeina per pirminę žievę, išskirdama fermentus, kurie sunaikina žievės ląsteles savo kelyje. Ankstyvosiose vystymosi stadijose šis pradas sudaro šaknies dangtelį, viršūnines ir pirmines meristemas. Laikui bėgant, dėl parenchiminių ląstelių diferenciacijos į ksilemos ir floemo elementus, centriniai šoninių ir tėvų šaknų cilindrai yra tarpusavyje susiję.
Vienatūriams pirminė šaknies struktūra išsaugoma visą gyvenimą, o antrinių audinių neatsiranda. Su amžiumi ląstelių dydis gali padidėti, ląstelių membranos sustorėti, tačiau bendras šaknies struktūros „planas“ išlieka nepakitęs.
Gimnazijose ir dviskilčiuose vyksta antriniai pakitimai, susiformuoja antrinė šaknies struktūra. Antrinės šaknies struktūros susidarymas visų pirma yra susijęs su kambiumo veikla. Kambiumas susidaro dalijant tas progamines ląsteles, kurios lieka nediferencijuotos, tarp pirminės floemos ir pirminės ksilemos. Tada per dviračio ląstelės, esančios ne ksilemo projekcijose, taip pat demonstruoja kambinį aktyvumą.
Kambis antrines ksilemo ląsteles nusodina į centrą, o antrines floemines - periferijoje. Taip susidaro laidžios sijos. Jų susidaro tiek, kiek buvo pirminės ksilemos spindulių. Spaudžiant augančius antrinius audinius, pirminės floemos ląstelės išsilygina. Pirminė ksilema išlaiko centrinę padėtį ir yra matoma kaip žvaigždutė. Laikui bėgant pirminė ksilema taip pat išnyks.
Esant antrinei šaknies struktūrai, pirminiai audiniai neveikia kaip laidūs elementai. Medžiagų srautas abiem kryptimis atliekamas per antrinius audinius.
Kambiumas, kilęs iš dviračio, sudaro plačius parenchiminius spindulius, esančius tarp atvirų užstato pluoštų. Tai yra pagrindiniai šerdies spinduliai. Jie užtikrina fiziologinį ryšį tarp centrinės šaknies dalies ir pirminės žievės. Vėliau galima nutiesti siauresnius antrinius šerdies spindulius, jungiančius antrinę ksilemą ir floemą.
Antriniai pokyčiai vyksta ne tik centriniame cilindre. Dėl padidėjusio laidžių audinių storio pirminė žievė stumiama į išorę, plyšta ir kartu su endoderma pašalinama.
Išorinėje dviračio dalyje atsiranda helogenas, kuris nusėda į išorę, o į vidų - daugiasluoksnis. Taigi, antrinės struktūros dviskilčių šaknys yra padengtos peridermu.
Daugiametėse šaknyse hellogenas atsiranda šaknies gilumoje. Tada šaknis, kaip ir stiebas, sudaro ritmą.
Svarbus šaknies antrinio augimo ekologinis aspektas yra skirtingų tos pačios rūšies augalų šaknų gebėjimas natūraliai augti. Kai šaknys liečiasi viena su kita, jos susijungia per antrinį augimą. Susikaupus šaknims, dirigavimo sistemos tęstinumas nustatomas tarp abiejų įgimtų partnerių. Dėl šaknų kaupimosi daugelis medžių yra tarpusavyje susiję, o jei dalis jų nukirsta, kelmai ilgai išlieka gyvi. Šaknų kaupimasis skatina greitą infekcijos perdavimą iš vieno medžio į kitą.
Šaknų diferenciacija šaknų sistemose pasireiškia įvairiomis formomis. Skirstymas į pagrindines, šonines ir atsitiktines šaknis yra labai svarbus.
Šaknys, atsirandančios aukštesnių sporų augalų viršūninėse meristemose, yra suskirstytos į specialią grupę. Manoma, kad jie yra archajiškiausia šaknų grupė ir išlaikė didžiausią panašumą su pirminėmis šaknimis, kurios buvo vienos išsišakojančios antžeminių ir požeminių ašių sistemos dalis. Šiai grupei priklausančios šaknys vadinamos apyzigeninėmis.
Remiantis šaknų sistemų atsiradimo seka evoliucijos procese ir priklausant didelėms sisteminėms grupėms, trys šaknų sistemų evoliucinis lygis.
I. Aukštesnių sporų augalų šaknų sistemas vaizduoja išskirtinai apygeninių šaknų sistemos, viena po kitos pakeičiančios viena kitą, o tai yra pagrindinis šaknų sistemų atsinaujinimo mechanizmas. Šakojimas yra viršūninis arba šoninis, ir gali būti, kad abi šakos rūšys atsirado nepriklausomai viena nuo kitos. Nėra siauros specializacijos metamorfozių ir šaknų.
P. Gimnastikos šaknų sistema išsiskiria tuo, kad yra pagrindinė šaknis, kuri gali būti siejama su sėklos ir dvipolio embriono su embrionine šaknimi atsiradimu. Į atnaujinimo mechanizmą įtrauktos ne tik šoninės pavaldžių kategorijų šaknys, bet ir rizogeninės atsitiktinės šaknys (t. Y. Atsitiktinės šaknys ant šaknų, atsitiktinės šaknys ant ūglių nesulaukė pastebimo vystymosi gimnazijose). Specializuotos ir metamorfizuotos šaknys yra retos.
Sh. Šakniavaisių šaknų sistemai būdingas stiprus atsitiktinių šaknų vystymasis, taip pat siauros specializacijos metamorfozės ir šaknys. Atnaujinimo mechanizmai yra įvairūs ir efektyvūs.
Šaknies sistemų metamorfozė
Šaknies sistemų metamorfozės yra anatominių ir morfologinių charakterių rinkinys, sukurtas filogenetiškai ir paveldimai fiksuotas ir atsiradęs dėl fiziologinių funkcijų pokyčių. Metamorfozė (metamorfozė, graikų kalba - transformacija) yra genetinių lygių pokyčių, sukeliančių organo, organų sistemos ar viso organizmo morfologinius ir funkcinius pertvarkymus, rezultatas.
I. Metamorfozės, susijusios su netipinių funkcijų atlikimu:
1) kvėpavimo šaknys;
2) epifitų oro šaknys;
3) šakotosios šaknys;
4) palaikyti šaknis.
P. Metamorfozės, susijusios su hiperfunkcija:
1) šakniavaisiai;
2) šaknies kūgiai;
3) šaknies mazgeliai;
4) mikorizė.
Šakniavaisiai būdingi dvimečiams dviskilčiams augalams. Tai įprastas pavadinimas. Jie vystosi iš vegetatyvinių organų ir neturi nieko bendra su vaisiais. Apatinė stiebo dalis ir šaknis dalyvauja formuojant šakniavaisį.
Pirmaisiais gyvenimo metais augalas auga, formuojasi šakniavaisiai. Antraisiais metais susidaro gėlės, vaisiai, sėklos. Tai reiškia, kad šakniavaisiai yra medžiagų tiekimo organas, leidžiantis augalui perkelti ramybės periodą ir visiškai vystytis kitais metais.
Šakniavaisių rūšys:
1. Monokambinis.
Pradinė šaknis yra su diarchiniu ašiniu cilindru. Šakniavaisio formavimasis prasideda antriniais pokyčiais, susijusiais su vieno kambrinio žiedo sukūrimu ir veikimu bei tolesne parenchimizacija.
Mes žinome, kad kambi antrinę ksilemą nusodina į vidų, o antrinį floemą - į išorę. Anatominiai skirtumai yra susiję su saugojimo audinio vystymosi vieta. Jei floemoje vyksta stiprus parenchimos vystymasis, susidaro floeminis šakniavaisių tipas - morkų rūšis. Jei ksilemoje - ksilemo tipas - ropės tipas.
2. Polikambinis šakniavaisių tipas.
Uždėtas vienas tipiškas kambijaus žiedas, kiti yra papildomi periciklinės kilmės. Jų gali būti iki 25. Šie papildomi kambio žiedai sudaro atvirus užpakalinius kraujagyslių ryšulius. Kuo žiedas yra toliau nuo pirmojo kambrinio žiedo, tuo daugiau saugojimo parenchimos jis sukuria. Polikambinių šakniavaisių pavyzdys yra burokėliai.
Vieno ir daugiakamžio šakniavaisiuose pirminė žievė anksti nudžiūsta ir nudžiūsta. Dengiantis audinys yra peridermis.
Taigi bendras visų šakniavaisių bruožas yra tai, kad yra daug saugojimo parenchimos, prasiskverbiančios iš laidžių audinių. Glaudus ryšys tarp laidžių audinių ir saugojimo audinių pasiekiamas įvairiais antrinio augimo modifikacijomis.
Kai kuriuose augaluose šakniavaisiai formuojami iš šoninių ir atsitiktinių šaknų šaknies kūgių pavidalu. Vienaląsčių (lubka) šakniniai kūgiai yra nekambijiniai, turi pirminę struktūrą; dviskilčiai - kambiniai, turi antrinę struktūrą. Pavyzdys: Phloem tipo dahlia šaknies kūgiai.
Šakniniai kūgiai per visą paviršių sudaro atsitiktines šaknis, o viršutinėje dalyje - atsitiktinius pumpurus, todėl yra vegetatyvinio dauginimo organai.
Yra žinoma, kad augalų augimą dažniausiai riboja nepakankamas fiksuoto azoto kiekis. Azotas stabilios N 2 molekulės pavidalu sudaro 80% atmosferos. Prieš fiksavimą (N 2 redukciją į amoniako NH 3) ši molekulė turi būti suskaldyta. Azoto fiksavimą atlieka kai kurios laisvai gyvenančios bakterijos, kurios sunaudoja dirvožemio organines medžiagas. Kai kurios bakterijos nusėda ant aukštesnių augalų šaknų. Augalas „pritraukia“ bakterijas organinėmis šaknų išskyromis. O mikroorganizmai savo ruožtu aprūpina augalą fiksuotu azotu. Ši abipusiai naudinga dviejų organizmų asociacija vadinama simbioze. Dažniausiai šaknys sudaro asociacijas su Rhizobium genties azotą fiksuojančiomis bakterijomis. Į simbiozę su bakterijomis sugeba įsikurti daugiau nei 60 dviskilčių ir vienaląsčių augalų šeimų. Kiekviena augalų rūšis turi savo simbiotinį Rhizobium. Abipusis augalų ir bakterijų atpažinimas atliekamas prie konkrečios bakterijos prijungiant specialų baltymą - lektiną, esantį trichoblastų paviršiuje. Šaknies plaukai deformuojasi veikiami bakterijų išskiriamų augimo hormonų. Bakterijos patenka į šias susuktas ląsteles. Ląstelių viduje dauginasi bakterijos ir susidaro infekcinis siūlas, uždengtas dantenų apvalkalu. Šis siūlas giliai įsiskverbia į šaknį. Šio bakterijų įsiskverbimo rezultatas yra stiprus šaknies mezoderminių ląstelių dauginimasis, dėl kurio susidaro mazgeliai. Išorinė mazgelio dalis ilgą laiką išlieka meristematinė zona, o vidinėje mazgelio dalyje yra bakterijų. Pačiam bakterijų audiniui atstovauja mažos ląstelės su klampia citoplazma ir daugybė bakterijų. Bakterinis audinys yra apsuptas laidžių ryšulių, sujungtų su centriniu šaknies cilindru. Tie. yra transporto sistema, skirta keistis maistinėmis medžiagomis tarp bakterijų ir augalų. Azotą fiksuojančios bakterijos teigiamai veikia augalų mitybą azotu ir dirvožemio derlingumą.
Fiziologinį šaknies aktyvumą, susijusį su augalo aprūpinimu vandeniu ir mineralais, galima sustiprinti simbioziniu ryšiu su konkrečiu grybeliu. Tokia abipusiai naudinga šaknų ir grybų simbiozė yra mikorizė. Mikorizė yra žinoma daugelyje kraujagyslių augalų. Tik kelios žydinčių augalų šeimos jo nesudaro arba retai formuojasi, pavyzdžiui, kopūstai, viksvos.
Mikoriziniai grybai vaidina pagrindinį vaidmenį mineralinių kraujagyslių augalų mityboje. Svarbus mikorizės vaidmuo absorbuojant ir perduodant fosforą.
Pervežant abipusiai (fosfatai juda iš grybelių į augalus, o angliavandeniai-priešinga kryptimi), apoplastinėje zonoje tarp augalo ir grybelio membranos prarandama ir absorbuojama.
Mikorizės grybai naudoja P šaltinius, kurių augalai paprastai neturi. Mikorizė taip pat skatina Zn, Mn, Cu įsisavinimą augaluose. Šie dirvožemio elementai yra silpnai mobilūs, todėl aplink šaknų plaukus greitai susidaro išeikvotos zonos. Mikorizės grybų hifų tinklas tęsiasi kelis centimetrus nuo kiekvienos šaknies ir taip padidina šaknų absorbcijos paviršių.
Skiriami šie mikorizės tipai:
1) ektomikorizė: grybelis storu dangteliu dengia visą šaknies galiuką ir įsiskverbia į tarpląstelines erdves;
2) endomikorizė: grybelis įsiskverbia į ląsteles.
Ektomikorizė būdinga tam tikroms medžių ir krūmų šeimoms (pušis, gluosnis, bukas). Ektomikorizė susidaro ant jaunų, dar neužkimštų šaknų. Ektomikorizės šaknys yra trumpos, šakotos ir atrodo patinusios.
Tokiose šaknyse slopinamas šaknies plaukelių vystymasis ir galima sumažinti viršūninės meristemos ir šaknies dangtelio tūrį. Šaknies plaukų funkciją atlieka hifai.
Endomikorizinės šaknys savo forma yra panašios į įprastas, tačiau nuo jų skiriasi tamsesne spalva. Grybeliniai hifai prasiskverbia į šaknies žievės ląsteles, kur sudaro pūsleles (patinimus) ir arbūzus (į medžius panašias šakas). Todėl endomikorozė dažnai vadinama vezikuline-arbuskuline. Esant tam tikroms sąlygoms, ląstelės gali „suvirškinti“ grybelinius hifus. Šaknų plaukai ant šaknų yra išsaugoti. Tai yra labiausiai paplitęs mikorizės tipas. Pasitaiko maždaug 80% visų kraujagyslių augalų, būtent žolinių augalų, įskaitant javus; krūmai, medžiai (obelys, klevai ...). Pavyzdžiui, jauname tuopos augale susidaro vezikulinė-arbuskulinė endomikorizė, o senstant ji muilina ektomikorizę.
Erikoidinė-arbutoidinė mikorizė būdinga viržiams Ericaceae. Grybelis aplink šaknį sudaro galingą hifų grupę, o plonos šoninės hifos prasiskverbia į šaknies žievės ląsteles. Šie mikoriziniai grybai žymiai sumažina dirvožemio toksiškumą viržiams, leisdami šiems augalams įsikurti rūgščiame, maistinių medžiagų neturinčiame dirvožemyje.
Yra dar viena rūšis - orchidėjų endomikoriza. Orchidėjų sėklos gamtoje dygsta tik esant atitinkamiems grybams.
Apskritai, esant natūralioms sąlygoms, mikorizės asociacijos yra taisyklė, o ne išimtis.
Seniausių kraujagyslių augalų liekanų tyrimas parodė, kad endomikorozė jose pasireiškė ne rečiau nei šiuolaikiniuose augaluose. Tai leido manyti, kad mikorizės atsiradimas evoliucijos procese gali būti vienas iš veiksnių, leidusių augalams apgyvendinti žemę. Atsižvelgiant į prastą dirvožemio išsivystymą šios gyvenvietės metu, galima daryti prielaidą, kad mikoriziniai grybai buvo augalų mineralinės mitybos pagrindas. Šiuolaikiniai pionieriniai augalai, turintys mikorizę, dažniau išgyvena skurdžiose dirvose.
Augalai, turintys mikotrofinę mitybą, yra atsparesni sunkiųjų metalų buvimui aplinkinėje technogeninėje aplinkoje.
Keletas žodžių apie atogrąžų miškų augalų šaknis. Dažniau nei kuokštinės, į lentas panašios atraminės šaknys. Šaknys primena lentas, o viršutiniai galai atsiremia į medį. Kai sulenktos šaknys yra atsitiktinės šaknys, atsirandančios ant kamieno ir augančios žemyn, lentų šaknų formavimasis yra susijęs su šoninėmis šaknimis, augančiomis tiesiai po dirvos paviršiumi. Šoninėse šaknyse išsivysto vertikalios ataugos, trikampio skerspjūvio, besiribojančios su medžio kamienu.
Lentos šaknų mediena dažnai yra kietesnė nei likusi to paties medžio mediena, ir dažnai jie yra padengti plonu žievės sluoksniu. Sprendžiant iš augimo žiedų, matomų šių šaknų skerspjūvyje, iš pradžių tai yra normalios, suapvalintos šaknys, bet vėliau jos labai stipriai sustorėja viršutinėje pusėje. Lentų šaknų skaičius medyje svyruoja nuo 1 iki 10, tačiau daugeliu atvejų jų yra trys ar daugiau. Tropikuose į lentas panašių šaknų susidarymas ypač pastebimas pelkėtuose miškuose augančiuose medžiuose ir prastai nusausintame dirvožemyje. Lentų šaknys dažnai naudojamos pramonės reikmėms. Pavyzdžiui, į stalą panašios šaknys iš Indonezijoje augančio Koompassia excelsa medžio naudojamos valgomojo stalams gaminti.
PABĖGIMAS
Ūgliai, kaip ir šaknis, yra pagrindinis augalo vegetatyvinis organas. Šaudymas atlieka oro tiekimo funkciją, taip pat gali atlikti daugybę kitų funkcijų ir gali metamorfozuoti.
Nekintamą vegetatyvinį ūglį sudaro ašinė dalis - stiebas; lapai ir pumpurai. Pagrindinis išorinis bruožas, skiriantis ūglį nuo šaknies, yra ūglio lapiškumas.
Stiebo dalis, susijusi su šoninių organų (šakų, lapų ir kt.) Atsiradimu - mazgas. Stiebo dalis tarp gretimų mazgų - tarpvietės. Vidinis kampas tarp lapo ir stiebo - lapo sinusas. Susiformuoja kiekvienas pasikartojantis ūglio elementas (mazgas su lapu ir pumpuru lapų pažastyje ir tarpvietėje) metamer. Taigi pabėgimas susideda iš metamerų serijos, t.y. turi metamerinę struktūrą.
XX amžiaus 90 -ųjų pradžioje Amerikos mokslininkai, vietoj mazgo ir tarpinio mazgo sąvokų, analizavo augalo augimą ir diferenciaciją, buvo pasiūlyta ši koncepcija - stiebo elementas. Stiebo elementas yra stiebo dalis tarp lapų primordijos kraštų. Jis aptinkamas šaudymo viršūnėje, kur vis dar nėra tarpinio mazgo. Tie. naujoji „kamieninio elemento“ sąvoka atitinka „metamero“ sąvoką.
Ūglio viršūninės meristemos struktūra yra sudėtingesnė nei šaknis. Iš tiesų, ūglio viršūninė meristema ne tik sudaro pirminių audinių ląsteles, bet ir dalyvauja formuojant lapų primordijas ir pažasties pumpurus, kurie vėliau išsivysto į šoninius ūglius. Ši meristema nėra apsaugota į šaknies dangtelį panašią struktūrą.
Viršūnių tipai yra tokie patys kaip ir šaknyje: mono-pradinis, zoninis, daugiapakopis. Pakopų skaičius yra didesnis nei šaknyje.
Evoliuciškai primityvi mono-pradinė viršūnė. Šio tipo asiūklio viršūnių pavyzdys. Viena didelė, pailga viršūninė ląstelė yra stipriai vakuolizuota. Skiriasi lygiagrečiomis savo paviršiams kryptimis (išskyrus išorinį paviršių). Artimiausios išvestos ląstelės taip pat stipriai vakuolizuojamos, tačiau dalijantis atsiranda mažų ląstelių su tankiais protoplastais. Tokios ląstelės randamos išilgai stiebo viršūnės periferijos, kur dedami lapų pumpurai.
Daugumos žydinčių augalų ūglio vegetatyvinė viršūnė pastatyta kaip tunika - kūnas - dvi zonos, besiskiriančios ląstelių dalijimosi plokštumose. Tunika(lot. - apvalkalas) apima išorinius ląstelių sluoksnius, dalijančius antiklininius (statmenus paviršiui) ir užtikrinančius paviršiaus padidėjimą. Rėmas(lot. - kūnas) - centrinė ląstelių grupė po tunika, jos dalijasi į skirtingas plokštumas, sudarydamos pagrindinį ūglio tūrį. Kūnas ir kiekvienas tunikos sluoksnis turi savo inicialus. Tunikoje inicialai dedami centre išilgai ašies. Dvispalviai iš esmės turi dviejų sluoksnių tuniką, vienaląsčiai-vieno, rečiau dviejų sluoksnių.
Nors tunikos kūno sąvoka padeda suprasti ląstelių vietą ir augimą pačioje viršūninėje meristemoje, skirtingų ląstelių zonų atpažinimas gretimose srityse padeda atskleisti meristemos indėlį į audinių ir organų diferenciaciją. šaudykloje. Viršūninėje meristemoje yra zonų, turinčių skirtingas citologines charakteristikas citologinis zonavimas.
Kūnas atitinka ląstelių sritį, vadinamą centrinių motinos ląstelių zona.Šios zonos ląstelės yra labai vakuolizuotos - tai savybė, susijusi su palyginti mažu mitozinio aktyvumo greičiu. Centrinių motinos ląstelių zoną supa periferinė meristema, iš dalies kylanti iš tunikos, iš dalies iš kūno. Protoderminis histogenas yra kilęs iš išorinio tunikos sluoksnio. Periferinė meristema gaunama iš šoninių viršūninių inicialų darinių ir iš centrinių motinos ląstelių. Medulinė meristema susidaro dėl padalijimų išilgai centrinių motinos ląstelių periferijos sluoksnyje, vadinamame pereinamoji zona. Periferinės zonos ląstelės turi tankius protoplastus, todėl turi didelį mitozinį aktyvumą.
Tam tikrose vietose, kuriose yra ypač didelis aktyvumas, klojami lapų gumbai (arba pirminiai gumbai).
Naujo lapų primordiumo atsiradimas yra susijęs su ląstelių dalijimosi krypties pasikeitimu, prieš kurį pasikeičia periferinių citoplazmos sluoksnių mikrotubulų orientacija. Osmosinis slėgis taip pat veikia primordiumo augimą.
Aktyvaus augimo laikotarpiu ūglio viršūninė meristema taip greitai suformuoja lapų primordijas, kad iš pradžių mazgai ir tarpvietės nesiskiria. Palaipsniui plotai tarp lapų pakopų pradeda augti, išsitempti ir įgauti tarpdančių išvaizdą. Tuo pačiu metu lapų tvirtinimo sritys yra suformuotos į mazgus.
Žemiau, kur atsiranda lapų užuomazgos, pastebima vakuolizacija. Tai rodo pirminės žievės ir žievės diferenciacijos pradžią. Stipri vakuolizacija pirminėje žievėje ir šerdyje rodo, kad yra meristematinė zona, vadinama liekamąja meristema. Atrodo, kad tai būtų viršūninės meristemos liekana, išsaugota tarp labiau diferencijuotų audinių. Žemiau lapų primordiumo vietos liekamojoje meristemoje išilginiais padalijimais, be tolesnio ląstelių augimo pločio, susidaro šiek tiek pailgos pirmojo prokambo ląstelės.
Kituose vystymosi etapuose vis daugiau prokambialinių krypčių diferencijuojasi likusioje meristemoje. Po to, kai susidaro visi tam tikro lygio kraujagyslių ryšuliai, liekamoji meristema diferencijuojasi į tarpfaskulinę parenchimą. Mazguose dalis likusios meristemos tampa lapų tarpo parenchima.
Prokambį galima kloti į tvirtą žiedą arba į atskiras gijas. Jei prokambi atsiranda kaip kietas žiedas, jis susidaro bekaroliukas (ne karoliukas) tipo stiebo struktūra. Jei prokambija klojama atskiromis gijomis, ji susidaro sijos tipo stiebo konstrukcija. Todėl stiebe pirminiai laidieji audiniai išsidėstę įvairiai: pirmuoju atveju - kietas žiedas, antruoju - atskirais laidžiais ryšuliais, atskirti pagrindinės parenchimos sekcijomis - meduliariniais spinduliais.
Antraeiliuose kraujagyslių ryšuliuose pirminė floema atsiranda išorinėje prokambialinės sruogos dalyje, pirminė ksilema - vidinėje. Vėlesnis floemo diferenciacija įvyksta centripetaliai: nauji floemo elementai atsiranda arčiau stiebo centro. Ksilemas skiriasi priešinga kryptimi - išcentrinis.
Visa stela arba centrinė stiebo dalis susideda iš laidžių audinių, šerdies (kartais ji sugriūna) ir dviračio. Pirminė žievė yra už dviračio ribų. Pirminė žievė (ir ją sudaro periferinė meristema) apima chlorenchimą, kartais sekrecinius elementus. Išorinis periferinės meristemos sluoksnis nesudaro egzodermos, kaip šaknyje, bet diferencijuojasi į kollenchimą, kuris yra sruogų arba kieto žiedo pavidalu (priklausomai nuo stiebo struktūros: ryšulio arba ne) pluoštas). Vidinis pirminės žievės sluoksnis yra endoderma. Tačiau stiebe jis neatlieka reguliavimo funkcijos, kaip ir šaknyje. Krakmolo grūdeliai nusėda endodermoje, todėl jie vadinami krakmolinga makštimi. Kartais jame yra kalcio oksalato kristalų.
Periciklas greitai praranda savo meristematinį aktyvumą ir yra paverčiamas sklerenchimu, kuris yra ištisiniame sluoksnyje arba virvelėse, priklausomai nuo stiebo struktūros tipo.
Šerdį vaizduoja plonasienės parenchiminės ląstelės, kuriose kaupiasi atsarginės maistinės medžiagos. Yra idioblastų ląstelių, pripildytų kristalų, gleivių ir kt. Periferinė šerdies dalis vadinama perimerulinė zona(Graikų peri - aplink, medulla - šerdis). Perimerulinės zonos ląstelės yra mažesnės ir ilgą laiką išlaiko savo gyvybinę veiklą.
Vienaląsčiams pradinis stiebo struktūros tipas yra kekių tipas, t.y. prokambiumą dengia virvelės. Tačiau antrinė meristema - kambis - niekada neatsiranda, todėl nėra antrinių laidžių audinių. Net ir sustorėjus, stiebą formuoja tik pirminiai audiniai. Storas vienaląsčių stiebas (delnai, alavijas) išsivysto dėl pagrindinės parenchimos ląstelių dalijimosi ir tempimo. Šis augimas vadinamas difuziniu antriniu augimu.
Vienaląsčių paukščių pirminė žievė ir centrinis cilindras nėra atskirti aiškia siena, kaip ir dviskilčiai, o stela artėja prie epidermio.
Vienaląsčiams būdingas difuzinis stiebo sandaros tipas, kai uždari laidūs ryšuliai yra išsibarstę po visą stiebą. Pailgėjant tarpuplaučiams, pirminiai flioemos ir ksilemo elementai (protofloemas ir protoksilemas) yra ištempiami ir sunaikinami, dėl to ryšulio ksilemo pusėje susidaro didelė oro ertmė. Todėl, atliekant laboratorinius tyrimus, kai mes žiūrėjome į ryšulių tipus, nuolat ruošiant kukurūzų kotelio skerspjūvį, buvo matyti, kad subrendęs laidus ryšulėlis apima du plačius metaksilemo indus, oro ertmę ir metafloem. Kiekvienas ryšulis yra uždarytas sklerenchiminių ląstelių apvalkalu.
Vienaląsčiams stiebo šerdis dažnai miršta, o jo vietoje susidaro ertmė. Toks tuščiaviduris stiebas su aiškiai apibrėžtais mazgais vadinamas šiaudų. Solominai būdingas stiprus mechaninio audinio vystymasis, kuris sudaro žiedą stiebo periferijoje. Subrendusiuose augaluose chlorenchyma sunaikinama, o šiaudai įgauna aukso geltonos spalvos, būdingos sklerenchimai. Apskritai vienaląsčiams būdingas stiprus stiebo sklerifikacija, kai iki 30% visų audinių yra sklerenchimoje ir gali būti dalinis lignifikavimas gyvų parenchiminių ląstelių membranų.
Dvilapių stiebuose antrinis augimas yra kambio veiklos rezultatas. Kambis atsiranda pirmųjų ūglių augimo metų pabaigoje ir skatina stiebo sustorėjimą. Kambis iš dalies susidaro iš prokambio, esančio kraujagyslių ryšuliuose, ir iš dalies iš tarpląstelinės parenchimos. Priklausomai nuo susidarymo vietos, kambio sekcijos vadinamos ryšuliu ir tarpfaskuliniu kambiumu. Būsimos kamieninės ląstelės interfundus zonose nustatomos daug anksčiau nei prasideda kambio veikla. Kambis pirmiausia susidaro ląstelėse, esančiose greta ryšulio, o paskui plinta giliai į medulinį spindulį. Dėl to tarpfundas ir pluošto kambis sudaro ištisinį kamieno žiedą stiebe.
Yra trys antrinio augimo tipai:
1) tipas Kirkazon Aristolochia.
Inicialas yra stiebo struktūros ryšulio tipas. Pirminės laidžiosios sijos yra atskirtos plačiomis šerdimis. Pluošto kambis sudaro antrinius laidžius audinius, tarpfaskulinis kambis sudaro spinduliuotės parenchimą. Ši pluošto struktūra yra išsaugota.
2) saulėgrąžų Helianthus rūšis.
Ryšio tipo struktūra taip pat yra pradinė. Tačiau antriniai laidieji audiniai susidaro tiek iš ryšulio, tiek iš sąveikos. Antriniai laidieji ryšuliai, suformuoti tarpinio sijos kambiumo, skiriasi nuo pirminių mažesniu dydžiu ir tuo, kad virš ryšulio nėra mechaninio periciklinės kilmės audinio. Ilgai veikiant kambiui, antrinių ryšulių tampa vis daugiau. Jie auga, susilieja tarpusavyje, sudarydami ištisinį antrinių laidžių audinių žiedą.
3) Tilia liepų tipas.
Inicialas yra stiebo struktūros be kuokštų tipas. Paprastasis kambijos žiedas sudaro antrinius laidžius audinius. Tačiau kartas nuo karto kambis sudaro parenchimines ląsteles, kurios vėliau diferencijuojasi į antrinius spindulius.
Daugiamečių augalų stiebo struktūra
Medžių ir krūmų dviskilčiai, taip pat spygliuočiai, antriniai sustorėjimai tęsiasi daugelį metų, o kai kurių rūšių kamienai siekia kelis metrus. Sustorėjimai yra susiję su ilgai veikiančio kambiumo ir, iš dalies, helogeno, dviejų antrinių šoninių meristemų, veikla. Topografiškai daugiamečio sumedėjusio augalo stiebą galima suskirstyti į tris pagrindines dalis: žievę, medieną ir šerdį. Riba tarp žievės ir medžio eina palei kambiumą.
Kambio ląstelės, priešingai nei tipiškos meristematinės ląstelės, visada yra stipriai vakuolizuotos. Kambio ląstelių citoplazmos būklė keičiasi sezoniškai.
Kambialinėje zonoje arba kambio žiede yra dvi kamieninių ląstelių formos: verpstės formos ląstelės (inicialai), pailgos vertikalia kryptimi, ir sferinės ląstelės, pailgos horizontalia kryptimi - spindulių inicialai. Jei fusiforminės ląstelės yra išdėstytos taisyklingomis horizontaliomis eilėmis, kambis vadinamas pakopiniu. Jei fusiforminių ląstelių išdėstyme tokios tvarkos nėra, kambis yra pakopinis.
Ilgųjų ūdų kambiumas sukelia ilgamedinę medieną ir antrinę ūdą. Nepakopinis kambis gamina daugiapakopę medieną ir nepakopinę antrinę floemą. Evoliuciniu požiūriu pakopiniai miškai ir floemai laikomi labiau specializuotais nei daugiapakopiai.
Sustorėjus stiebui, padidėja ir kambi sluoksnių perimetras. Tai įmanoma dėl kamieninių ląstelių dalijimosi radialine kryptimi.
Tarp kamieninių ląstelių išskiriamas vidurinis pradinis sluoksnis. Jo ląstelės dėl dalijimosi sudaro darinius vidine, o paskui išorine kryptimi. Per tą patį laikotarpį į vidų nusėda 2-3 ląstelės, o išorėje-tik viena ląstelė. Dėl to susidaro daugiau medienos nei masto.
Spinduliuotės ląstelės (inicialai) sudaro medulinius parenchiminius spindulius: pirminius, kurie tęsiasi nuo smegenų ir žievės; o antriniai šerdies spinduliai yra trumpesni, nes atsiranda iš vėliau atsiradusių spindulių ląstelių.
Pagrindinės sijos gali būti vienos eilės ir kelių eilučių; vienalytis (t. y. susidedantis iš tų pačių ląstelių) ir nevienalytis (spindulys apima skirtingos struktūros ląsteles).
Pagrindinių spindulių reikšmė : tai maistinių medžiagų (krakmolo, lipidų) saugojimo vieta, kuri atlieka tam tikrą vaidmenį ramybės laikotarpiu (pavyzdžiui, žiemą).
Šerdies spinduliai užtikrina radialinį vandens ir mineralinių druskų pernešimą iš ksilemos išilgai apoplasto į kambiumą ir antrinę floemą, o priešinga kryptimi - išilgai maistinių medžiagų simplasto.
Ir, pagaliau, dujų mainai vyksta palei tarpląstelines erdves.
Mediena
Mediena susidaro iš kambio ir auga išcentrine kryptimi. Kadangi mūsų zonoje kambis veikia periodiškai: jis pradeda veikti ankstyvą pavasarį ir nustoja veikti vasaros pabaigoje, tada medyje matomas koncentriškas sluoksniavimasis. Pavasarį, vegetacijos pradžioje, susidaro ankstyva mediena. Jo indai yra platūs, plonasieniai, gali gabenti daug vandens. Vanduo reikalingas augimui pradėti, ypač norint padidinti naujų ląstelių dydį (pvz., Besivystančių lapų ląstelės). Vėlyvoje medienoje yra mažiau indų. Jie yra siauresni, storesni. Tais pačiais metais perėjimas nuo ankstyvosios iki vėlyvosios medienos gali būti laipsniškas. Tačiau vienų metų vėlyvosios medienos siena su kitų metų ankstyvuoju medžiu visada yra aštri ir aiškiai matoma skerspjūvyje. Rezultatas yra metinis augimas (metinis žiedas arba metinis sluoksnis).
Pagal indų vietą metiniame (metiniame) augime išskiriamos šios medienos rūšys.
Jei indai daugiausia yra ankstyvojoje medienoje, bet vėlesniuose jų nėra arba dedami tik maži indai, pvz., Ąžuolo ar uosio mediena, tai žiedinė kraujagyslių mediena.
Jei indai tolygiai pasiskirsto per visą metinį augimą, nors jų skersmuo vėlyvoje medienoje mažėja, tai yra išsklaidyta kraujagyslių mediena. Rasta beržo, klevo, tuopos.
Tarp šių medienos rūšių taip pat yra pereinamųjų formų.
Kadangi mediena atlieka laidžiosios, mechaninės ir saugojimo funkcijas, ji, be ksilemo elementų, taip pat apima mechaninį audinį sklerenchimos pavidalu ir saugojimą pagrindinės parenchimos pavidalu. Visų anatominių elementų struktūra, jų skaičius, išdėstymas suteikia skirtingų augalų medienai specifinių ypatybių, būdingų tam tikrai taksonominei grupei.
Pavyzdžiui, labiau evoliuciškai pažengę augalai turi specializuotą mechaninį audinį (sklerenchimą). Evoliuciškai primityvūs dvikojai (magnolijos) neturi medienos pluošto. Tracheidai šiuo atveju atlieka mechaninį vaidmenį.
Kai kurių augalų medienoje yra pereinamosios formos nuo tracheidų iki libriforminių (libriform yra specializuoti mechaniniai medienos elementai, susidedantys iš prosenchiminių ląstelių, nukreiptų į galus su storomis lignifikuotomis membranomis). Taip pat randama „Cloisonné libriform“. Jo originali fusiforminė ląstelė su storomis vertikaliomis membranomis yra padalinta į atskiras trumpas gyvas ląsteles. Tokie histologiniai elementai morfologiškai ir funkciškai artėja prie sumedėjusios parenchimos.
Medinė parenchima atlieka saugojimo ir iš dalies laidų vaidmenį. Parenchimos ląstelės yra gyvos. Medžio parenchima yra horizontalioje ir vertikalioje eilutėse. Horizontalios eilutės sudaro pagrindines sijas. Mes jau apsvarstėme jų savybes. Vertikalios eilutės sudaro pačią medinę parenchimą.
Jei medžio parenchima yra išsibarstę per visą metinį augimą, tai yra difuzinė parenchima. Būdinga liepų, ąžuolo, kriaušių medienai.
Jei sumedėjusi parenchima yra aplink indus, tai yra paratrachealinė parenchima. Pelenuose, klevo medienoje.
Ant išorinės metinio augimo ribos yra galinė parenchima. Pavyzdys: gluosnio, maumedžio, magnolijos mediena.
Jo struktūros ypatumas yra kraujagyslių nebuvimas. Trachėjos elementai nėra perforuoti ir juos vaizduoja tracheidos, kurios atlieka ir laidų, ir mechaninį vaidmenį.
Spygliuočių tracheidoms būdingos didelės ribotos poros su toru. Tracheidos ilgos su smailiais galais. Ankstyvosios tracheidės yra didelės, plonasienės. Vėlyvosios tracheidos yra radialiai suplotos, storomis sienomis.
Matome, kad gimnazijos ir dviskilčių mediena turi didelių struktūrinių skirtumų. Todėl jis suskirstytas į dvi pagrindines grupes. Gimnastikos mediena klasifikuojama kaip minkšta, o dviskilčių mediena - kaip kieta mediena. Šie terminai „kietas“ ir „minkštas“ nebūtinai apsiriboja tankio ir kietumo laipsniu. Skirtumai, kaip jau minėjome, yra susiję su struktūrinėmis ypatybėmis.
Reaktyvi mediena susidaro nuožulniais arba išlenktais kamienais. Jo susidarymą lemia šakų ir kamienų polinkis atsispirti įtempiams, atsirandantiems dėl pasvirusios padėties, t.y. jo susidarymas yra susijęs su šių augalų dalių tiesinimo procesu.
Svarbiausi reaktyviosios medienos vystymosi veiksniai yra gravitacinės jėgos ir endogeninio augimo stimuliatorių pasiskirstymas.
Reaktyvi mediena skiriasi nuo įprastos medienos tiek anatomiškai, tiek chemiškai. Reaktyviosios medienos ląstelės yra labiau linkusios lignifikuotis ir sustorėti. Spygliuočiuose jis yra tankesnis ir tamsesnis nei aplinkiniai audiniai. Jo tracheidės yra trumpesnės nei įprastos medienos.
Stiebo medienos radioaktyvumas mažėja nuo išorinių sluoksnių iki šerdies. Nuo bagažinės pagrindo iki viršaus šiek tiek padidėja medienos užterštumas.
Pakankamai didelis radioaktyviųjų medžiagų kiekis žievėje ir ūgliuose rodo jų išorinę oro taršą.
Žievė
Šis terminas apima visus audinius, esančius už kambiumo ribų. Tai jau antrinė pluta, nes susidaro antrinė meristema - kambis (priešingai nei pirminis, kuris susidaro diferencijuojant pirminės meristemos ląsteles).
Antrinė žievė apima antrinę floemą, sklerenchimą (pluoštus ir akmenuotas ląsteles) ir pagrindinę parenchimą. Mechaninių elementų rinkinys vadinamas kietas niekšas laidūs elementai ir pagrindinė parenchima - minkštas bastas.
Antrinėje žievėje yra sekrecinės ląstelės, dervos kanalai. Pagrindinė antrinės žievės funkcija yra nešioti plastines medžiagas, taip pat apsauginė funkcija.
Antrinė floema sudaro daug mažesnę kamieno dalį nei antrinė ksilema. Taip yra dėl to, kad, pirma, kambis sudaro mažiau floemo nei ksilemas, ir, antra, senas, neveikiantis floemas palaipsniui suglamžomas.
Dėl padidėjusio ašinio organo apskritimo parenchiminiai spinduliai plečiasi ir primena trikampį, kurio viršūnė nukreipta į kambiumą. Tokia parenchima vadinama išsiplėtimu (lot. Dilatatio - išsiplėtimas). Šis parenchimos išdėstymas apsaugo liemenį nuo plyšimo augant stiebui storio.
Spygliuočiuose antrinė floema turi paprastesnę struktūrą. Gimnastikos sėklų sluoksnyje histologiniai elementai yra tolygiai pasiskirstę ir, žinoma, yra dervos kanalų.
Visi audiniai, esantys už helogeno (tai reiškia vidinį kamštienos kamibį) ribų, kai kamštienos ląstelės suberinizuojasi, nustoja gauti vandens ir mineralų. Visi šie peridermiai su žievės liekanomis (negyvosios parenchimos ir floemos ląstelės) sudaro išorinę (išorinę) žievę. Gyva žievės dalis, esanti tarp kambiumo ir vidinio helogeno sluoksnio, vadinama vidine žieve.
Taigi, remdamiesi savo žiniomis iš temos „Audiniai“, ištyrėme stiebo kaip ūglio ašinio organo anatominę struktūrą.
Dabar apsvarstykime ūglio šoninio organo struktūrą - lapą.
LAPAS
Lapas yra pagrindinis aukštesniųjų augalų fotosintezės organas. Lapo struktūra ir funkcijos yra glaudžiai susijusios.
Iš bendros fotosintezės reakcijų lygties:
СО 2 + Н 2 О chl-l → hv (СН 2 О) n + О 2
galime daryti išvadą, kad 1) lapams reikia CO 2 ir vandens šaltinio; 2) lapai turi būti pritaikyti susigerti saulės energija jie turi turėti chlorofilo; 3) O 2 bus išleistas kaip vienas iš reakcijos produktų; 4) angliavandeniai turi būti laikomi arba vežami į kitas augalo dalis. Sąrašas yra labai specializuota įstaiga, atitinkanti visus šiuos reikalavimus.
Visa lapo, kaip vegetatyvinio organo, evoliucija vyko prisitaikymo prie vystymosi keliu geriausia naudoti Sveta.
Pagal mikrofilminę evoliucijos liniją lapas laikomas rinofito tipo archajinių augalų telomo atauga ir išlaiko ašinės organo struktūros struktūrą.
Išilgai makrofilinės evoliucijos linijos, lapas atsirado dėl išlyginimo, telomos buvo išdėstytos vienoje plokštumoje ir sujungtos į vieną visumą. Tuo pačiu metu buvo prarastas ilgalaikio viršūninio augimo ir išsišakojimo gebėjimas. Kokios morfologinės ir anatominės lapo savybės atsirado dėl ilgos evoliucijos?
Suaugusį lapą paprastai sudaro lapų ašmenys ir lapkočiai. Žiedlapis- siaura į stiebą panaši lapo dalis tarp lapo ašmenų ir šaudymo mazgo, kurios pagalba lapas orientuojasi erdvėje ir yra palankiausias šviesos atžvilgiu. Per lapkočius atliekamas ryšys tarp lapų ašmenų ir stiebo. Lapai gerai išvystę laidžius ir mechaninius audinius. Nukritus lapams, lapkočio lapeliuose susidaro atskyrimo sluoksnis.
Žemiausia lapo dalis, sujungta su stiebu, vadinama lapo pagrindas. Lapo pagrindas gali įgauti įvairių formų. Dažniausiai jis atrodo kaip nedidelis sustorėjimas ir vadinamas lakštų pagalvėlė. Dažniau lapo apačioje yra suporuoti įvairių formų ir dydžių šoniniai ataugos - stipules. Stipules gali būti lapo formos ir tarnauti kaip papildomas asimiliacinis paviršius. Stupulai gali būti aštrūs ir atlikti augalų apsaugos funkcijas.
Yra pleiskanojančių, plėvelių kuokelių. Evoliuciniu požiūriu stubreliai sumažėja, o evoliuciškai jaunuose augaluose jie anksti nukrinta arba jų visai nėra.
Kai kurių javų, salierų (skėčio) lapų pagrindas auga ir sudaro uždarą arba atvirą vamzdelį - lapų apvalkalą. Lapų apvalkalas prisideda prie ilgalaikio tarpslankstelinės meristemos išsaugojimo ir yra papildoma priemonė šaudymui.
Pagrindinė lapo dalis yra lapų ašmenys. Dėl netolygaus viršutinės ir apatinės lapo pusių apšvietimo plokštė turi dorsoventralinę struktūrą, t.y. viršutinės lapo pusės, ventralinės, struktūra skiriasi nuo apatinės pusės, nugaros struktūros. Viršutinė lapo pusė vadinama ventraline, nes inkstuose ši pusė yra vidinė ir pasukta į ūglio ašį. Apatinė, nugarinė, pumpuro lapo pusė yra pasukta į išorę nuo augalo. Taigi lapas su dorsoventraline simetrija yra dvišalis arba dvišalis arba dvipusis. Būdinga dviskiltėms.
Lapas su radialine simetrija yra lygiavertis. Tokie sustorėję cilindriniai lapai randami niekšuose. Vienpusiai arba izoliuoti vienpusiai lapai būdingi javams ir svogūnams.
Apsvarstykite lapų ašmenų anatominė struktūra.
Lapas, kaip ir šaknis bei stiebas, susideda iš vientisų, laidžių ir pagrindinių audinių. Kadangi lapas paprastai neturi antrinio augimo (išskyrus nedidelį lapkočių ir didelių venų augimą); jis išlaiko epidermį kaip dengiamąjį audinį.
Stomatai daugiausia yra apatinėje lapo pusėje. Dviskilčiuose lapuose stomatai išsibarstę po visą lapų paviršių be matomos tvarkos. Vienaląsčių ir spygliuočių lapuose jie pasiskirsto eilėmis, lygiagrečiomis išilginei lapo ašiai. Kai kuriuose grūduose epidermyje yra motorinių ar motorinių ląstelių. Jie yra didesni už įprastas epidermio ląsteles. Vakuolė užima beveik visą ląstelę. Praradus turgorą, šios ląstelės susitraukia ir prisideda prie lapų sulankstymo ar garbanojimo.
Dauguma pagrindinis lapų ašmenų audinys yra mezofilas. Mezofilas yra diferencijuotas. Palisade mezofilas yra viršutinėje plokštės pusėje, kempininis - iki apatinės. Tokios struktūros lapai yra bifaliniai arba dorsoventraliniai.
Jei palisade mezofilas yra abiejose lapo pusėse, lapas yra vienpusis arba izoliuotas.
Grūdų mezofilyje nėra aiškios diferenciacijos į palisadą ir kempinę chlorenchimą. Mezofilo ląstelės yra radialiai aplink ryšulius.
Lapo kraujagyslių ryšuliai vadinami gyslomis, o šakų sistema - venomis venavimas. Laidžių sijų išdėstymo lapų plokštėse ypatumai lemia skirtingi tipai venavimas, kuris gali būti dvejopas, lygiagretus, lankinis, pirštas ir plunksninis.
Skiriamos atviros (dichotominės) ir uždarosios vencijos, atsižvelgiant į tai, kaip tarpusavyje susijusios aukštesnių šakų eilių gyslos. Esant dichotominėms venoms, venos išsišakoja dichotomiškai, išsišakoja aštriu kampu ir pasiekia lapo ašmenų kraštą, niekur nesusilieja su gretimomis venomis. Uždarame venavime dėl anastomozių (kryžminimo) mažos venos sudaro tinklą, į kurį prasiskverbia visa plokštelė arba didžioji jos dalis.
Lapai su pirštų ir plunksnų venomis, kurie daugiausia yra dviskilčiai, turi vidutinę, didžiausią gyslą. Veną sudaro pirminė ksilema ir pirminė floema, atsirandanti iš prokambiumo ir sujungta į užstatą. Tarp ksilemo ir floemo dažnai susidaro kambis, tačiau jis neveikia. Todėl ksilemas ir floemas yra pagrindiniai.
Ryšiai sudaro ištisinę sistemą lape, prijungtą prie laidžiosios stiebo sistemos. Todėl ksilemas visada yra orientuotas į morfologiškai viršutinę lapo pusę, o floema - į apatinę morfologinę pusę.
Vidurinis kraštas yra sujungtas į šoną su šoninėmis venomis, kurių dydis yra mažesnis. Kiekvienas iš jų yra susijęs su dar mažesniais, o jie, savo ruožtu, yra suskirstyti į dar mažesnius ir t. Išsišakojusių užsakymų skaičius dviskilčiais svyruoja nuo 2 iki 5 ar daugiau. Mažiausios šakos sudaro ląsteles, kurių viduje yra uždarytos mažos mezofilo sritys - areola.
Atviroje dichotominėje venų sistemoje uždarų ląstelių nėra.
Vienaląsčių lapų (lygiagrečiai) venose išilginės gyslos yra sujungtos mažesnėmis gyslomis - komisiniais ryšuliais, kurie išdėstyti paprastų skersinių tiltų pavidalu. Laidžių audinių skaičius mažose venose palaipsniui mažėja. Ryšių galuose ksilemo elementai dažnai tęsiasi didesnį atstumą nei floeminiai. Ksilemą venų galuose paprastai sudaro trumpi trachėjos elementai, floemą - trumpi siauri sieto elementai ir didelės palydovinės ląstelės.
Mažus kraujagyslių ryšulius, lokalizuotus mezofilyje, supa vienas ar keli kompaktiškai sulankstytų ląstelių sluoksniai, sudarantys ryšulio apvalkalą. Pluošto apvalkalai gali būti parenchiminiai arba sklerenchiminiai. Kai kurioms rūšims stebimas ryšulių apvalkalų ląstelių membranų suberinizavimas, o tai rodo, kad apvalkalo ląstelės gali veikti kaip endodermas.
Kollenchima ir sklerenchima yra didelėse venose iš vienos ar abiejų pusių. Su didelėmis venomis susijęs audinys pakyla virš lapo paviršiaus ir sudaro iškyšas. Audinių sritis, esanti tarp venų iškyšų, vadinama „tarpšonkauline zona“ (lotyniškas iškyšos pavadinimas „costa“ - šonkaulis).
Rūšiuose su storais lapais mezofilyje yra įvairių sklereidų, kurie veikia kaip tarpikliai.
Grūdų lapams būdingas stiprus sklerenchimos vystymasis.
Gimnastikos lapų struktūra yra mažiau įvairi nei angiospermų. Panagrinėkime, naudodamiesi pušų spyglių pavyzdžiu, kaip prisitaikymas prie žemos temperatūros žiemą ir gebėjimas gyventi keletą metų atsispindi anatominėje struktūroje. Adata turi storų sienelių epidermį su galinga odele ir giliai palaidotomis stomatomis. Kai kurių spygliuočių (Cupressaceae) šoninės ląstelės turi kutikulinį keterą, kuri iškyla ant stomatų apsauginių ląstelių. Stomata yra išdėstyta vertikaliomis eilėmis iš visų adatų pusių. Po epidermiu yra storos sienelės hipodermos ląstelės su iš dalies lignifikuotomis membranomis. Mezofilas yra sulankstytas. Jame yra dervos ištraukų. Adatų centre yra laidūs ryšuliai. Paprastai du, retai vienas. Šie ryšuliai yra sujungti sklerenchiminiu lygintuvu. Ryšulius supa perpylimo audinys, sudarytas iš tracheidų ir parenchiminių ląstelių. Perpylimo audinys (lot. Transfusio - perpylimas) yra susijęs su vandens ir maistinių medžiagų transportavimu tarp kraujagyslių ryšulių ir mezofilo.
Perpylimo audinį supa storasienis endodermas. Endodermas dažnai turi Caspari diržus ankstyvose vystymosi stadijose, o vėlesniuose - suberino sluoksnį. Subrendusios būklės endodermos ląstelės turi antrines lignifikuotas membranas.
Anatominė plokštės struktūra atskleidžia didelį plastiškumą. Struktūra kinta priklausomai nuo augalo egzistavimo sąlygų ir lapų išdėstymo ant augalo.
Mes jau pažymėjome vieną iš šiuolaikinės botanikos vystymosi krypčių - ekologinę augalų anatomiją - ir pasakėme, kad būtent gyvas ląstelės turinys - protoplastas - pirmiausia reaguoja į aplinkos sąlygų pokyčius. Taigi šiaurinių augalų ląstelės yra mažiau vakuolinės ir turi didelius chloroplastus. Stromoje yra daug krakmolo grūdų. Chloroplastų tylakoidinė sistema yra mažiau išvystyta nei chloroplastų vidutinio klimato platumų augaluose. Šiaurės augaluose mitochondrijų skaičius yra 2–2,5 karto didesnis. EPR ir Golgi aparatai yra labiau išvystyti. Tokios mezofilo ląstelių submikroskopinės struktūros ypatybės šiauriniuose augaluose apibūdinamos kaip svarbios prisitaikančios savybės, prisidedančios prie augalų išlikimo atšiauriomis klimato sąlygomis.
Augalai, prisitaikę prie drėgmės trūkumo, turi storą ląstelių membraną, ypač epidermyje, šaltu oru. Stipriai sustorėjusios išorinės didžiųjų ląstelių epidermio membranos užima pusę ląstelių ertmės. Virš epidermio yra storas odelių sluoksnis. Iš odelės paviršiaus ji padengta storomis vaško žvyneliais, kurie lemia melsvą lapų atspalvį.
Stomatos yra palaidotos epidermio plaukų išstumtose įdubose.
Sultingi augalai rodo, kad yra vandens kaupiantis audinys.
Įdomūs prietaisai drėgmei sugauti iš minkštųjų kvinojos lapų (Afrika). Viršutinė ir apatinė lapo pusės yra padengtos ištisiniu pūslelių pavidalo plaukų sluoksniu, išdėstytu keliomis eilėmis. Šio sluoksnio storis žymiai viršija mezofilo storį ir apsaugo gyvus audinius nuo išdžiūvimo. Plaukai gali sulaikyti drėgmę ore naktį ir pagerinti atšiaurų augalo vandens režimą.
Mūsų plintančios miško zonos kvinojos lapai ir stiebas yra padengti daugybe pūslelių panašių plaukų, kurie sudaro miltelių žydėjimo įspūdį, tačiau jie nesudaro tokio tankaus, kelių eilučių sluoksnio, kaip minkštoje kvinojoje.
Kaip pavyzdį palyginus dviejų rūšių kvinojos lapus, galima pastebėti, kad artimų taksonominių vienetų sisteminiai charakteriai, kai jie įsikuria skirtingomis ekologinėmis sąlygomis, įgyja nevienodą vystymąsi ir yra prisitaikantys.
Atogrąžų miškų augalų (tokių kaip monstera) lapai turi perforuotus lapų ašmenis ir ilgus lapkočius. Vandens perteklius visada lengvai nutekėja. Stomata yra atvira. Palisadinis audinys su didelėmis oro kišenėmis.
Be temperatūros, drėgmės, dar vienas svarbus abiotinis veiksnys yra šviesa, į kurią augalas reaguoja su struktūriniais pokyčiais.
Lapai, kurie vystosi tiesioginiuose saulės spinduliuose, yra mažesni už šešėlinius lapus, bet storesni už šešėlinius lapus dėl stipraus palisade mezofilo vystymosi.
Taigi evoliucijos eigoje fiksuoti prisitaikymai prie aplinkos sąlygų yra susiję su kiekybiniais skirtumais, tačiau nekeičia organizavimo principų.
Lapų ontogenezė
Lapai dedami mažų gumbų pavidalu kaip šoniniai viršūninės meristemos ataugos. Formuojant lapų gumbą, dalyvauja ne tik tunika, bet ir kūno ląstelės. Lapų primordiumas auga nuo pradinės projekcijos į viršų kūgio arba adatos formos ataugos pavidalu. Netrukus lapų užuomazgos supa viršūninę meristemą, apsaugodamos ją tiek mechaniškai, tiek karščiu, kurį jie išskiria kvėpavimo metu.
Ankstyvas lapų augimas paprastai skirstomas į viršūninį ir ribinį (ribinį) augimą. Pirmasis yra susijęs su primordiumo pailgėjimu, antrasis - su šoniniu augimu, dėl kurio susidaro dvi plokštelės dalys. Atitinkamai, augantis primordiumas turi viršūninę meristemą viršūnėje ir dvi kraštines meristemas šoninėje ašyje. Pirminis viršūninis augimas yra trumpas. Po viršūnių ir sluoksnių pakraščių augimo seka tarpslankstelinis augimas.
Sudėtingame lape kraštinės meristemos veikla yra lokalizuota atskiruose centruose, iš kurių kiekvienas sudaro atskirą lapą.
Lapų ašmenys vystosi greičiau nei lapkočio augimas, kuris vėliau susidaro tarpkaulinio augimo metu.
Jei riba tarp tarpakarinės ir ribinės zonos pagrindo susidaro susiaurėjimas, tada lapkočiai išsivysto. Didžiąją lapkočio dalį sudaro tarpkaulinė zona. Jei toks susiaurėjimas (ties riba tarp tarpląstelinių ir kraštinių zonų pagrindo) nesusiformuoja, išsivysto sėdimas lapas.
Vienaląsčių augalų lapų primordiume kraštinės meristemos beveik nesusiformuoja, o tarpląstelinis augimas būdingas ilgą laiką. Taigi pagrindinė lapų ašmenų forma yra linijinė.
Ląstelių dalijimosi ir pailgėjimo greičio skirtumai skirtinguose sluoksnių sluoksniuose lemia daugelio tarpląstelinių erdvių susidarymą ir tipišką mezofilo struktūrą.
Kitų audinių vystymasis koreliuoja su laidžių audinių vystymusi. Prokambis vienu metu yra izoliuotas lape ir gretimoje augimo kūgio dalyje, sudarant ištisinę giją. Jis sudaro pirminį kraujagyslių ryšulį, kuris yra ištisinis ir bendras lapams ir stiebams.
Lapo ontogenezė liudija apie bendrą lapo ir stiebo prigimtį kaip į vieną visumą - ūglį.
Nuo to momento, kai pumpuras išsiskleidžia, prasideda ekstrarenalinė lapų vystymosi fazė. Tokiu atveju lapų paviršius padidėja kelis šimtus, net tūkstančius kartų. Paviršiaus augimas pasiekiamas padalijus daugumą lapų ląstelių ir ištempiant jas į ilgį ir plotį.
Lapų gyvenimo trukmė priklauso nuo genetinių ir klimato veiksnių. Vidutinio klimato lapuočiuose ir krūmuose ekstrarenaliniai lapai gyvena 4-5 mėnesius. Spygliuočių lapai gyvena nuo dvejų iki šešerių metų ar ilgiau. Bet kokiu atveju lapų gyvenimo trukmė yra žymiai trumpesnė nei ašinių augalo organų.
Welwitschia mirabilis auga uolėtose pietvakarių Afrikos dykumose. Šis nykštukinis medis turi ilgą šaknį, storą ir trumpą kamieną (iki 50 cm aukščio ir iki 1 m storio) ir du didelius lapus, kurie išlieka visą gyvenimą (iki 2000 metų). Lapai siekia 2-3 metrus ir nuolat auga prie pagrindo, miršta viršuje. Beveik vienintelis Velvichia drėgmės šaltinis yra tankus rūkas, kurio drėgmę šis augalas sugeria per daugybę stomatų abiejose lapo pusėse (22 200 stomatų 1 cm 2).
Aktyvus lapų atskyrimas nuo šakų nepažeidžiant jo gyvų audinių vadinamas abscissija. Sezoninis medžių lapų kritimas yra reakcijos į dienos ilgio pokyčius rezultatas. Kritę lapai rudenį žymiai sumažina augalo garinimo paviršių, kuris būtinas esant fiziologiniam drėgmės trūkumui rudenį ir žiemą. Dėl nukritusių lapų sumažėja šakų lūžimo pavojus nuo sniego sunkumo.
Daugumoje lapų atskiriantis sluoksnis susidaro ontogenezės metu. Šioje srityje sumažėja mechaninio audinio kiekis. Prieš nusileidimą trachėjos elementuose susidaro teles, o sietuose - kalozė. Krentant, fermentinis ląstelių membranų sunaikinimas, dėl kurio ląstelės atsiskiria viena nuo kitos. Pokyčiai membranose apima vidutinio sluoksnio cementinio gebėjimo praradimą (iš dalies dėl to, kad iš jo pašalinamas kalcis), pačių celiuliozės membranų hidrolizę ir sklerifikuotų trachėjos elementų plyšimą.
Apsauginis sluoksnis arba randas susidaro dėl to, kad membranose ir tarpląstelinėse erdvėse nusėda apsauginės medžiagos, tokios kaip suberinas ar žaizdos guma. Sumedėjusiems augalams apsauginį sluoksnį pakeičia peridermė, kuri klojama po apsauginis sluoksnis ir tampa ištisinis su likusio stiebo peridermė.
Lapų kritimas nebūtinai yra susijęs su ląstelių membranų ištirpimu. Mechaninį atskyrimą be išankstinių cheminių pokyčių galima pastebėti, kai spyglius numeta nuo eglės.
Nukritus lapams, lieka lapų randai su jų ryšulių randais. Ryšių randai- tai sulaužytų laidžių sijų galai, kurie nuo lapų pėdsakų iki lapų lapkočių nukrito.
Lapų pėdsakai- tai šakos nuo stiebo laidžios sistemos iki lapų. Lapų takas driekiasi nuo jo santakos vietos su stiebo ryšuliu iki išėjimo į lapą. Vienas lapas gali turėti vieną ar daugiau lapų žymių.
Kadangi stiebas ir lapas yra filogenetiškai paplitę, ir nėra esminio skirtumo tarp lapų pėdsakų ir stiebo kuokštų. Susiję terminai turi aprašomąją, topografinę reikšmę.
Mazgo srityje, kur lapų pėdsakai nukrypsta nuo stiebo centrinio laidžio cilindro link lapo pagrindo, cilindre susidaro parenchiminė zona - lapų spraga arba lapų tarpas. Lapų pėdsakų ir lapų spragų skaičius skirtinguose augaluose skiriasi.
Pagrindinis angiospermų mazgų tipas yra mazgas, turintis tris ar daugiau spragų. Pagrindinei angiospermų evoliucijos krypčiai būdingas spragų skaičiaus sumažėjimas.
Tarpai, šerdys, tarpinės sijos, laidžioji sistema ir periciklas, esantys laidžiosios sistemos periferijoje, sudaro vidinę ašinės augalo dalies (stiebo ir šaknies) šerdį arba stela.
Stelų klasifikacija grindžiama santykiniu laidžių ir nelaidžių audinių padėtimi ašiniuose organuose pradiniame vystymosi etape. Paprasčiausio tipo stelose, kurios laikomos filogenetiškai primityviausiomis, laidus audinys sudaro ištisinę koloną, o flioemas supa ksilemą. Tai yra protostelas. Tokia stiebo organizacija yra biologiškai netobula, nes kontaktinis paviršius tarp laidžių ir apatinių audinių yra mažas.
Tolesnė stelos evoliucija sekė laidaus audinio kontaktinio paviršiaus su pagrindiniais padidinimo keliu. Tai buvo pasiekta dviem būdais. Vienu atveju stela formuoja gilias ataugas žievės link ir turi žvaigždės formos skerspjūvį. Tai aktinosteles.
Antrasis, biologiškai perspektyvesnis vystymosi kelias yra parenchiminės šerdies atsiradimas stelės centre. Tai yra sifono stela.
Glaudesnis ryšys tarp laidžių ir pagrindinių audinių atsiranda vystantis lapams. Kaip daugiau lapų, tuo labiau stela yra prisotinta pagrindine parenchima. Stelė įgauna tinklo (graikų tinklo - diktato) išvaizdą. Tai yra diktostelė.
Šiuolaikinių žydinčių augalų stela yra dar labiau parenchiminė. Stele, kuri yra sruogų ir tarpinių sijų laidumo sistema, yra eustela (graikų eu - gera).
Vienaląsčių lapų pėdsakai pasiekia stiebo centrą, tada nukrypsta į periferiją. Todėl stiebo skerspjūvyje ryšuliai atrodo išsibarstę be jokios tvarkos. Ataktos iš graikų kalbos. netvarkingas. Iš čia kilęs ir vienašakės stelos pavadinimas - ataktostela. Jam būdingas didelis parenchimizacijos laipsnis, kai kiekvienas lapų takas "



































- Dydis: 2 MB
- Skaidrių skaičius: 36
PRISTATYMO APRAŠYMAS ŠAKNIŲ IR ŠAKNIŲ SISTEMOS 1. Funkcijos ir evoliucija skaidrėmis
ŠAKNIŲ IR ŠAKNIŲ SISTEMOS 1. Šaknies funkcijos ir evoliucinė kilmė. 2. Pirminė šaknies struktūra. 3. Antriniai šaknų pokyčiai. 4. Šoninių ir atsitiktinių šaknų formavimas. Šaknų sistemos. 5. Šaknų specializacija ir modifikavimas.
Šaknis yra ašinis organas, turintis radialinę simetriją ir neribotą laiką augantis dėl viršūninės meristemos veiklos. Lapai niekada neatsiranda ant šaknies, o viršūninė meristema visada yra uždengta dangteliu. Pagrindinė šaknies funkcija yra absorbuoti vandenį ir mineralus, tai yra aprūpinti augalą dirvožemio mityba. Be įvardytos pagrindinės funkcijos, šaknys atlieka ir kitas funkcijas: stiprina augalą dirvožemyje, leidžia augti vertikaliai ir nešti ūglius aukštyn; šaknyse vyksta antrinė įvairių medžiagų (amino rūgščių, alkaloidų, fitohormonų ir kt.) sintezė; saugojimo medžiagos gali būti nusodintos šaknyse; šaknys sąveikauja su kitų augalų šaknimis, dirvožemio mikroorganizmais ir grybais.
Šaknys atsirado iš rinofitų kūnų, pasklidusių dirvos paviršiuje. Evoliucijos eigoje kai kurios šių kūnų šakos pradėjo gilintis į dirvą ir sukėlė šaknis.
Šaknys pritaikytos geresnei dirvos mitybai. Šaknų atsiradimą lydėjo gilus visos jų struktūros pertvarkymas. Juose atsirado specializuoti audiniai. Medžiagų absorbavimo iš dirvožemio funkciją pradėjo atlikti jaunos šaknų galūnės. Jie palaiko gyvas ląsteles paviršiuje. Šios ląstelės sudarė svarbiausią funkcinį šaknies audinį - rizodermą.
Medžiagų absorbavimo iš dirvožemio funkciją pradėjo atlikti jaunos šaknų galūnės. Jie palaiko gyvas ląsteles paviršiuje. Šios ląstelės sudarė svarbiausią funkcinį šaknies audinį - rizodermą. Be to, evoliucijos procese padidėjo sugeriantis šaknies paviršius dėl trijų veiksnių: 1) gausaus išsišakojimo ir daugybės siurbimo galų susidarymo; 2) nuolatinis šaknų augimas ir siurbimo galų perkėlimas į naujas dirvos vietas; 3) šaknų plaukų formavimas.
Kadangi šaknys auga tankiame dirvožemyje, jo viršūninė meristema turi būti apsaugota. Viršūninės meristemos apsaugą nuo pažeidimų užtikrino šaknies dangtelio atsiradimas. Šaknų atsiradimą lėmė didėjantis klimato sausumas. Prasidėjus sausesniam klimatui sausumos augalai prisirišo prie vandens ir maistinių medžiagų ir sugeria iš jų substratą. Tačiau evoliucijos metu skirtingų augalų rūšių šaknų struktūra pasikeitė mažiau nei stiebo. Taip yra dėl to, kad sąlygos dirvožemio aplinkoje yra stabilesnės nei ore. Todėl šaknis laikomas „konservatyvesniu“ organu, nors pasirodė daug vėliau nei ūgliai. Šaknų formavimasis yra svarbi augalų aromorfozė. Jo dėka augalai sugebėjo įvaldyti sausesnius dirvožemius ir suformuoti didelius, kylančius ūglius.
Šaknies dangtelio amiloplasto reakcija į gravitaciją. Statolitų judėjimas vaidina svarbų vaidmenį kuriant fitohormonų gradientus, užtikrinančius vertikalų šaknų augimą.
Kviečių sodinuko (Triticum aestivum) šaknies sandara: A - šaknies sandaros schema; B - rizoderminių ir eksoderminių ląstelių diferenciacija. 1 - laidumo zona, 2 - siurbimo zona, 3 - ištempimo zona, 4 - padalijimo zona, 5 - šaknų plaukai, 6 - šaknies dangtelis.
Šaknies skerspjūvis (a - vienalapis, b - dviskiltis augalas)
Pirminė žievė kyla iš pralaidumo. Pagrindinę jo masę sudaro gyvos parenchiminės ląstelės su plonomis membranomis. Tarp jų suformuota tarpląstelinių erdvių sistema, pailga išilgai šaknies ašies. Dujos (CO 2) cirkuliuoja per tarpląstelines erdves. Dujos yra būtinos intensyviam metabolizmui žievės ir rizodermos ląstelėse palaikyti. Stiprus metabolizmas žievės ląstelėse yra būtinas daugeliui svarbių funkcijų: 1) žievės ląstelės aprūpina šakniastiebį plastikinėmis medžiagomis ir pačios dalyvauja medžiagų absorbcijoje ir laidume; 2) žievėje sintetinamos įvairios medžiagos, kurios vėliau perkeliamos į kitus audinius; 3) rezervinės medžiagos kaupiasi žievės ląstelėse; 4) žievėje dažnai yra grybų, formuojančių mikorizę, hifų.
Endodermos ląstelės išgyvena tris vystymosi etapus. Siurbimo zonoje endoderma yra pirmoje stadijoje. „Caspari“ diržai yra suformuoti jos ląstelių radialinių sienų viduryje. Caspari diržai blokuoja medžiagų judėjimą per ląstelių membranas, tai yra išilgai apoplasto. Antrąjį etapą galima stebėti šoninėje šaknų zonoje. Be to, su viduje ląstelės membranoje atsiranda plona suberino plokštelė. Tačiau endodermas vis dar gali laisvai praeiti tirpalus, nes jame lieka atskiros praėjimo ląstelės su plonomis sienelėmis. Trečiąjį endodermos vystymosi etapą galima pastebėti vienaląsčių šaknų zonoje. Vidinės ir radialinės jo ląstelių sienos yra labai sustorėjusios. Skersiniuose pjūviuose tokios ląstelės turi pasagos formą. Nėra prieigos langelių. Storasienis endodermas apsaugo laidųjį audinį ir padidina šaknies stiprumą.
„Caspari“ diržas yra neperšlampamas barjeras, verčiantis vandenį palikti apoplastą ir skubėti per endodermos ląstelių membranas į simplastą
Vanduo teka iš dirvos į šaknį: vanduo gali judėti palei apoplastą ir simplastą, kol pasieks endodermą. Tolesnis judėjimas palei apoplastą yra neįmanomas.
Įvairių tipų šaknies centrinio cilindro struktūra (pirminė struktūra): A-diarch, B-triarch, C-tetrarch, D-polyarch. A-B tipai būdingi dviskilčiams, D-daugeliui vienaląsčių. 1 - pirminės žievės plotas, 2 - pirminė floema, 3 - pirminė ksilema.
Galima išskirti keturis šaknies perėjimo iš pirminės struktūros į antrinę struktūrą etapus: 1) kambiumo atsiradimą tarp pirminės floemos ir ksilemos sričių; 2) hellogeno susidarymas per dviratį; 3) pirminės žievės išliejimas; 4) laidžių audinių radialinio išdėstymo pakeitimas užstatu.
Perėjimas iš pirminės šaknies struktūros į antrinę 1 - pirminė floema, 2 - pirminė ksilema, 3 - kambis, 4 - dviratis, 5 - endoderma, 6 - mezoderma, 7 - rizodermas, 8 - egzodermas, 9 - antrinė ksilema, 10 - antrinė floema, 11 - antrinė žievė, 12 - phellogen, 13 - fella.
Laidžiųjų audinių pirminio diferenciacijos schema žirnių šaknyse 1 - centripetalinė ksilemos diferenciacija, 2 - epidermis, 3 - pirminė žievė, 4 - endoderma, 5 - pirmą kartą diferencijuoti ksilemo elementai, 6 - nediferencijuoti ksilemo elementai, 7 - pirmą kartą diferencijuoti flioeminiai elementai, 9 - viršūninė meristema, 10 šaknų dangtelis.
Antriniai vienaląsčių šaknų pokyčiai. Didžioji dauguma vienaląsčių augalų išlaiko pirminę šaknies struktūrą iki gyvenimo pabaigos. Tačiau šiuo atveju daugelis šaknies elementų yra lignified. Medžių vienaląsčių (palmių, dracaena, jukos) šaknų žievėje iš parenchimos ląstelių arba iš dviračio atsiranda meristeminis sluoksnis. Iš jo susidaro uždarų laidžių sijų eilės. Po šios serijos laidų ryšulių pirminės žievės parenchimos periferinėje dalyje atsiranda naujas ugdomojo audinio sluoksnis. Šis meristemos sluoksnis sukuria naują laidžių ryšulių seriją. Taigi atsiranda šaknų sustorėjimas.
Atsitiktinės šaknys atsiranda ant įvairių augalų organų - ant stiebų, lapų ir šaknų. Atsitiktinės šaknys, atsiradusios ant stiebo, vadinamos kamieninėmis šaknimis, o tos, kurios atsirado ant šaknies, vadinamos kampinėmis. Šoninės ir atsitiktinės šaknys yra endogeninės kilmės, tai yra, jos dedamos į vidinius audinius.
Šoninės šaknies klojimas prasideda dalijant dviračio ląsteles. Šiuo atveju ant stelos paviršiaus susidaro meristematinis gumbelis. Po daugybės meristematinio gumbelio ląstelių dalijimosi atsiranda šoninė šaknis. Jis turi savo viršūninę meristemą ir dangtelį. Šoninis šaknies pumpuras auga, prasiskverbia per pirminę motinos šaknies žievę ir juda į išorę. Paprastai šoninės šaknys atsiranda prieš ksilemo elementus. Todėl jie yra išdėstyti taisyklingomis išilginėmis eilėmis išilgai šaknies. Jie atsiranda absorbcijos zonoje arba šiek tiek aukščiau. Šoninės šaknys klojamos akropetiškai, tai yra nuo šaknies pagrindo iki viršūnės.
Atsitiktinės šaknys paprastai dedamos audiniuose, galinčiuose atlikti meristematinį aktyvumą: pericikle, kambiume, hellogene. Endogeninis šoninių (ir atsitiktinių) šaknų formavimasis yra prisitaikantis. Jei šakojimasis vyktų viršūnėje, šaknies judėjimas dirvožemyje būtų sunkus.
Dichotominis šakojimasis Lycopodium clavatum 1 šaknų sistemoje - ploniausių šaknų izotominis dichotominis išsišakojimas
Mikorizė: A - ąžuolo ektotrofinė mikorizė, B, C - endotrofinė orchidų mikorizė. Lubinų šaknų mazgeliai
Šaknų funkcijos.Šaknis yra pagrindinis aukštesniojo augalo organas. Šaknų funkcijos yra šios:
Jie išsiurbia iš dirvožemio vandenį ir jame ištirpintas mineralines druskas, perneša juos stiebu, lapais ir reprodukciniais organais. Siurbimo funkciją atlieka įsiurbimo zonoje esantys šaknų plaukai (arba mikorizė).
Dėl didelio stiprumo augalas tvirtinamas dirvožemyje.
- Vandens, mineralinių druskų jonų ir fotosintezės produktų sąveikos metu sintezuojami pirminės ir antrinės metabolizmo produktai.
- Esant šaknų spaudimui ir transpiracijai, jonai vandeniniai tirpalai mineralinės ir organinės medžiagos išilgai šaknies ksilemo indų juda išilgai kylančios srovės į stiebą ir lapus.
- Maistinės medžiagos (krakmolas, inulinas ir kt.) Kaupiamos šaknyse.
- Šaknyse atliekama antrinių metabolitų (alkaloidų, hormonų ir kitų biologiškai aktyvių medžiagų) biosintezė.
- Šaknų meristematinėse zonose susintetintos augimo medžiagos (giberelinai ir kt.) Yra būtinos augalo oro dalių augimui ir vystymuisi.
- Dėl šaknų atliekama simbiozė su dirvožemio mikroorganizmais - bakterijomis ir grybais.
- Padedant šaknis, vegetatyvinis daugelio augalų dauginimas vyksta.
10. Kai kurios šaknys veikia kaip kvėpavimo organas (monstras, filodendras ir kt.).
11. Daugelio augalų šaknys veikia kaip „suglebusios“ šaknys (ficus banyan, pandanus ir kt.).
12. Šaknis gali metamorfozuoti (sustorėti pagrindinės šaknies formos „šaknys“ morkose, petražolėse ir kt.; Sustorėjus šoninėms ar atsitiktinėms šaknims, susidaro šakniagumbiai jurginuose, žemės riešutuose, žievelėse ir kt., Sutrumpėja šakniavaisiai. augalai).
Šaknis yra ašinis organas, paprastai cilindro formos, radialinės simetrijos, su geotropizmu. Jis auga tol, kol yra išsaugota viršūninė meristema, padengta šaknies dangteliu. Ant šaknies, skirtingai nuo ūglio, lapai niekada nesusidaro, bet, kaip ir ūgliai, šaknies šakos, formuojasi šaknų sistema.
Šaknų sistema yra vieno augalo šaknų kolekcija. Šaknies sistemos pobūdis priklauso nuo pagrindinių, šoninių ir atsitiktinių šaknų augimo santykio.
^ Šaknų ir šaknų sistemų tipai. Sėklos embrione visi augalo organai yra kūdikystės stadijoje. Pagrindinė arba pirmoji šaknis vystosi iš embriono šaknis. Pagrindinė šaknis yra visos šaknų sistemos centre, stiebas tarnauja kaip šaknies pratęsimas, ir kartu jie sudaro pirmos eilės ašį. Sritis, esanti pasienyje tarp pagrindinės šaknies ir stiebo, vadinama šaknies kaklelis.Šis perėjimas iš stiebo į šaknį pastebimas dėl skirtingo stiebo ir šaknies storio: stiebas yra storesnis už šaknį. Stiebo atkarpa nuo šaknies kaklelio iki pirmųjų embriono lapų - vadinami skilčialapiai poodinis kelias arba hipokotilas... Nuo pagrindinės šaknies šoninės eilės eilės šaknys tęsiasi į šonus. Tokia šaknų sistema vadinama esminis, daugelyje dviskilčių augalų jis gali išsišakoti. Šakota šaknų sistema yra šaknies sistemos tipas. Šoninis šaknies išsišakojimas pasižymi tuo, kad naujos šaknys yra nutiestos tam tikru atstumu nuo viršūnės ir yra suformuotos endogeniškai - ankstesnės tvarkos pirminės šaknies vidiniuose audiniuose dėl periciklinio aktyvumo. Kuo labiau šoninės šaknys nutolsta nuo pagrindinės šaknies, tuo didesnis augalų mitybos plotas, todėl yra specialių agronominių metodų, kurie padidina pagrindinės šaknies gebėjimą formuoti šonines šaknis, pavyzdžiui, suspaudžiant arba nardyti pagrindinės šaknies l / 3 jos ilgio. Po tam tikro laiko nardymo pagrindinė šaknis nustoja augti, o šoninės šaknys intensyviai auga.
Dviskilčių augalų pagrindinė šaknis, kaip taisyklė, išlieka visą gyvenimą, vienaląsčių embriono šaknis greitai miršta, pagrindinė šaknis nesivysto ir susidaro nuo ūglio pagrindo išlygosšaknys, kurios taip pat turi pirmosios, antrosios ir kt. užsakymų. Tokia šaknų sistema vadinama pluoštinės. Atsitiktinės šaknys, kaip ir šoninės, yra dedamos endogeniškai. Jie gali susidaryti ant stiebų ir lapų. Augalų gebėjimas plėtoti atsitiktines šaknis yra plačiai naudojamas augalų auginimui vegetatyvinio augalų dauginimo metu (dauginant stiebais ir lapų auginiais). Virš žemės stiebo auginiai daugina gluosnius, tuopas, klevus, juoduosius serbentus ir kt .; lapiniai auginiai - uzambara violetinė arba saintpaulia, kai kurios begonijų rūšys. Požeminiai modifikuotų ūglių (šakniastiebių) auginiai dauginami daug vaistiniai augalai, pavyzdžiui, pakalnutė, vaistinis kupenu ir kt. Kai kurie augalai formuoja daug atsitiktinių šaknų, kai užmuša apatinę stiebo dalį (bulvės, kopūstai, kukurūzai ir kt.), taip sukurdami papildomą mitybą.
Aukštesnių sporų augaluose (šarmuose, asiūkliuose, paparčiuose) pagrindinė šaknis visiškai neatsiranda, jie sudaro tik atsitiktines šaknis, besitęsiančias nuo šakniastiebio. Daugelyje dviskilčių žolinių šakniastiebių augalų dažnai išnyksta pagrindinė šaknis ir vyrauja atsitiktinių šaknų sistema, besitęsianti nuo šakniastiebių (sloga, dilgėlė, šliaužiantis vėdrynas ir kt.).
Kalbant apie įsiskverbimo į dirvą gylį, pirmoji vieta priklauso šaknies šaknų sistemai: rekordinis šaknų įsiskverbimo gylis, remiantis tam tikra informacija, siekia 120 m! Tačiau pluoštinė šaknų sistema, turinti daugiausia paviršines šaknis, prisideda prie velėnos dangos susidarymo ir apsaugo nuo dirvožemio erozijos.
Bendras šaknų sistemos šaknų ilgis yra skirtingas, kai kurios šaknys siekia kelias dešimtis ar net šimtus kilometrų. Pavyzdžiui, kviečių visų šaknų plaukelių ilgis siekia 20 km, o žieminių rugių bendras pirmosios, antrosios ir trečiosios eilės šaknų ilgis yra daugiau nei 180 km, o pridedant ketvirtos eilės šaknis - 623 km. Nepaisant to, kad šaknis auga visą savo gyvenimą, jos augimą riboja kitų augalų šaknų įtaka.
Šaknies sistemų išsivystymo laipsnis skirtinguose dirvožemiuose skirtingose natūraliose zonose yra nevienodas. Pavyzdžiui, smėlėtose dykumose, kur požeminis vanduo yra gilus, kai kurių augalų šaknys eina į 40 m ar didesnį gylį (Selino javai, ankštinių šeimos ankštiniai ir kt.). Pusiau dykumos trumpalaikiai augalai turi paviršutiniškasšaknų sistema, kuri yra pritaikyta greitam ankstyvo pavasario drėgmės įsisavinimui, kurio pakanka greitai praeiti visoms augalų vegetacijos fazėms. Molinguose, prastai gazuotuose taigos miškų zonos podzoliuose augalų šaknų sistema yra 90% susitelkusi paviršiniame dirvos sluoksnyje (10-15 cm), augalai turi „maitinančias šaknis“ (europines egles). Pavyzdžiui, Saksonija turi šaknis skirtingu laiku metų naudoja drėgmę iš skirtingų horizontų.
Labai svarbus šaknų sistemos pasiskirstymo veiksnys yra drėgmė. Šaknų kryptis yra didesnė drėgmė, tačiau vandenyje ir užmirkusiame dirvožemyje šaknys išsišakoja daug silpniau.
Šaknies sistemų išsivystymo laipsnis, šaknų įsiskverbimo gylis ir kitos šaknies plastinės savybės priklauso nuo išorinių sąlygų ir tuo pat metu yra paveldimai priskiriamos kiekvienai augalų rūšiai.
^ Jaunos šaknies zonos. Jaunoje šaknyje yra: 1) padalijimo zona, uždengta šaknies dangteliu; 2) ląstelių tempimo zona arba augimo zona; 3) siurbimo arba šaknų plaukų zona; 4) laidžioji zona.
^ Padalijimo zonažymi šaknies galiuką, padengtą išorėje šaknies dangtelis, apsaugo viršūninę arba viršūninę meristemą. Jaunas šaknies galiukas yra slidus liesti dėl ląstelių išskiriamų gleivių. Kai šaknis ilgėja, gleivės sumažina šaknies galo trintį prie dirvožemio. Pasak akademiko V.L. Komarov, šaknies dangtelis „kasa žemę“, jis apsaugo besidalijančias meristemos ląsteles nuo mechaninių pažeidimų, taip pat kontroliuoja teigiamas geotropizmas pati šaknis, tai yra, ji skatina šaknies augimą ir įsiskverbimą į dirvožemio gelmes. Šaknies dangtelį sudaro gyvos parenchiminės ląstelės, kuriose yra krakmolo grūdelių. Po dangčiu yra padalijimo zona arba šaknies kūgis, atstovaujamas pirminio ugdymo audinio (meristemos). Dėl aktyvaus šaknies viršūninės meristemos dalijimosi susidaro visos kitos šaknų zonos ir audiniai. Jaunos šaknies padalijimo zona yra tik 1 mm ilgio. Iš išorės jis skiriasi nuo kitų zonų geltona spalva.
^ Tempimo zona, arba augimo zona, kurio ilgis yra keli milimetrai, jis yra išoriškai skaidrus, susideda iš ląstelių, kurios praktiškai nesidalija, bet ištempiamos išilgai. Ląstelės padidėja, jose atsiranda vakuolių. Ląstelėms būdingas didelis turgoras. Tempimo zonoje atsiranda pirminių laidžių audinių diferenciacija ir pradeda formuotis nuolatiniai šaknies audiniai.
Virš tempimo zonos yra siurbimo zona. Jo ilgis yra 5–20 mm. Siurbimo zoną vaizduoja šaknų plaukai - epidermio ląstelių ataugos. Šaknies plaukelių pagalba vanduo ir druskos tirpalai absorbuojami iš dirvožemio. Kuo daugiau šaknų plaukelių, tuo didesnis šaknies įsiurbimo paviršius. 1 mm šaknies paviršiaus gali būti apie 400 šaknų plaukų. Šaknų plaukai yra trumpalaikiai, gyvena 10 - 20 dienų, po to jie miršta. Šaknies plaukų ilgis skirtinguose augaluose yra nuo 0,5 iki 1,0 cm. Jauni šaknų plaukai susidaro virš ištempimo zonos ir miršta virš absorbcijos zonos, todėl šaknies plaukų zona nuolat juda, kai šaknis auga ir augalas gali sugerti vandenį ir jame ištirpusias maistines medžiagas iš skirtingų dirvožemio horizontų.
Virš siurbimo zonos prasideda laidumo zona arba šoninių šaknų zona. Vanduo ir druskos tirpalai, kuriuos sugeria šaknis, per medienos indus pernešami į antžemines augalo dalis.
Tarp šaknų zonų nėra aštrių ribų, tačiau pastebimas laipsniškas perėjimas.
6. Šaknies metamorfozė. Jų biologinė reikšmė. Mikorizė. Dauguma tos pačios šaknų sistemos augalų yra labai skirtingi augimą ir čiulpia galūnės. Augimo galai paprastai yra galingesni, greitai pailgėja ir juda giliau į dirvą. Tempimo zona juose yra gerai išreikšta, o viršūninės meristemos energingai veikia. Čiulpimo galūnės, kurios gausiai atsiranda ant augimo šaknų, lėtai ilgėja, o jų viršūninės meristemos beveik nustoja veikti. Čiulpimo galai tarsi sustoja dirvoje ir intensyviai „čiulpia“.
Sumedėjusiuose augaluose, storas skeleto ir pusiau skeletasšaknys, ant kurių trumpalaikės šaknies skilčių... Šaknies skiltys, kurios nuolat keičia viena kitą, apima augimą ir čiulpimą.
Jei šaknys atlieka specialias funkcijas, keičiasi jų struktūra. Aštrus, paveldimas organo pakeitimas, kurį sukelia funkcijų pasikeitimas, vadinamas metamorfozė... Šaknų modifikacijos yra labai įvairios.
Daugelio augalų šaknys sudaro simbiozę su dirvožemio grybų hifomis, vadinamomis mikorizė(„Grybų šaknys“). Mikorizė susidaro ant čiulpiančių šaknų absorbcijos zonoje. Grybelinis komponentas leidžia šaknims lengviau gauti vandens ir mineralinių elementų iš dirvožemio, dažnai grybeliniai hifai pakeičia šaknų plaukus. Savo ruožtu grybas iš augalo gauna angliavandenių ir kitų maistinių medžiagų. Yra du pagrindiniai mikorizės tipai. Hifai ektotrofinis mikorizė sudaro dangtelį, apimantį šaknį iš išorės. Ektomikoriza yra plačiai paplitusi medžiuose ir krūmuose. Endotrofinis mikorizė dažniausiai pasitaiko žoliniuose augaluose. Endomikorozė yra šaknies viduje, hifos įvedamos į plutos parenchimos ląsteles. Mikotrofinė mityba yra labai paplitusi. Kai kurie augalai, pavyzdžiui, orchidėjos, apskritai negali egzistuoti be simbiozės su grybais.
Ant ankštinių augalų šaknų atsiranda specialios formacijos - mazgeliai kurioje įsikuria Rhizobium genties bakterijos. Šie mikroorganizmai sugeba įsisavinti atmosferos molekulinį azotą, paversdami jį surišta būsena. Kai kurias mazguose susintetintas medžiagas pasisavina augalai, bakterijos, savo ruožtu, naudoja medžiagas šaknyse. Ši simbiozė yra labai svarbi Žemdirbystė... Ankštiniuose augaluose gausu baltymų dėl papildomo azoto šaltinio. Jie aprūpina vertingais maisto ir pašarų produktais bei praturtina dirvą azotinėmis medžiagomis.
Labai paplitęs saugojimasšaknys. Paprastai jie yra sutirštėję ir labai parenchima. Stipriai sustorėjusios atsitiktinės šaknys vadinamos šaknies kūgiai, arba šakniagumbiai(jurginas, kai kurios orchidėjos). Daugelis, dažniau kas dvejus metus, augalų, turinčių šaknies šaknų sistemą, sukuria darinį, vadinamą šakninė daržovė... Formuojant šakniavaisį dalyvauja ir pagrindinė šaknis, ir apatinė stiebo dalis. Morkose beveik visą šakniavaisį sudaro šaknis, o ropės - šaknys sudaro tik žemiausią šakniavaisio dalį ( ryžių. 4.12).
Šaknys auginamų augalų atsirado dėl ilgo pasirinkimo. Šakniavaisiuose laikymo parenchima yra labai išvystyta, o mechaniniai audiniai išnyko. Morkose, petražolėse ir kitoje skėtinėje parenchimoje parenchima yra stipriai išsivysčiusi floemoje; ropėse, ridikuose ir kituose kryžmažiedžiuose, ksilemoje. Burokėliuose atsarginės medžiagos nusėda parenchimoje, susidariusios veikiant keliems papildomiems kambi sluoksniams ( ryžių. 4.12).
Susidaro daug svogūninių ir šakniastiebių augalų įtraukikliai, arba susitraukiantisšaknys ( ryžių. 4.13, 1). Jie gali sutrumpinti ir atitraukti ūglį į dirvą iki optimalaus gylio vasaros sausros metu arba žiemos šalčių... Atsitraukiančios šaknys turi sustorėjusius pagrindus su skersiniu tvirtumu.
Kvėpavimošaknys, arba pneumatoforai (ryžių. 4.13, 2) susidaro kai kuriuose atogrąžų sumedėjusiuose augaluose, gyvenančiuose deguonies trūkumo sąlygomis (taksodiumas arba pelkinis kiparisas; mangrovių augalai, gyvenantys pelkėtose vandenyno pakrančių pakrantėse). Pneumatoforai auga vertikaliai aukštyn ir išsikiša virš dirvos paviršiaus. Per šių šaknų skylių sistemą, susijusią su aerenchima, oras patenka į povandeninius organus.
Kai kuriuose augaluose, norint išlaikyti ūglius ore, papildomai palaikantisšaknys. Jie nutolsta nuo horizontalių lajos šakų ir, pasiekę dirvos paviršių, intensyviai šakojasi, virsdami stulpelinėmis formacijomis, palaikančiomis medžio vainiką ( stulpelinis bananų šaknys) ( ryžių. 4.15, 2). Sustingęsšaknys tęsiasi nuo apatinių stiebo dalių, suteikdamos stiebui stabilumo. Jie susidaro mangrovių krūmynų augaluose, augalų bendrijose, kurios vystosi atogrąžų vandenynuose, užliejamuose potvynio metu ( ryžių. 4.15, 3), taip pat kukurūzuose ( ryžių. 4.15, 1). Ficus guminės formos panašus į lentąšaknys. Skirtingai nuo stulpelinių ir kuokštinių, jie yra ne atsitiktinės kilmės, o šoninės šaknys.
Ryžiai. 4.15. ^ Palaikančios šaknys: 1 - dygliuotos kukurūzų šaknys; 2 - į stulpą panašios bananų medžio šaknys; 3 - šakotos rizoforos šaknys ( ir kt- potvynių zona; nuo- atoslūgio zona; dumblas- purvino dugno paviršius).
Pabėgimo koncepcija. Ūglio morfologinis skrodimas. Mazgai ir tarpusavio pleištai. Viršūninis ūglių augimas. Kūgio struktūra ir aktyvumas auga. Ūgliai yra stiebas, ant kurio yra lapai ir pumpurai.
Stiebo sritys, kuriose auga lapai vadinami mazgais.
Stiebų sekcijos tarp dviejų artimiausių mazgų vadinamas internodais.
Kampas tarp lapo ir aukščiau esančio mazgo vadinamas lapų pažastimi.
Lapo pažastyje susidaro pažastinis pumpuras. Pabėgimas susideda iš pasikartojančių sekcijų - metameriai.
Viename metamere yra tarpšonkaulis, mazgas, lapas ir pažasties pumpuras. Ūgliai yra kompleksas, susidedantis iš stiebo ir lapų. Pagrindinis ūglis dedamas į embrioną, kur jį vaizduoja inkstai. Pumpurą sudaro embrioninis stiebas - epikotilė, viršūninė meristema ir vienas ar daugiau lapų primordijų (lapų primordijos). Kai sėkla sudygsta, stiebas pailgėja. Naujos lapų primordijos vystosi iš viršūninės meristemos, nuo lapų primordija lapai vystosi, ir lapų pažastys susidaro inkstų primordijos. Šis kūrimo algoritmas gali būti daug kartų pakartotas formuojant augalų ūglių sistemą.
Suformuotame ūglyje išskiriami mazgai - ūglio dalis, kurioje lapas sujungtas su stiebu; tarpvietės - šaudymo dalis tarp mazgų, paprastai tai yra stiebo dalis; lapų sinusai - kampas tarp lapo ir kylančios stiebo dalies.
Pumpurai taip pat yra šaudymo dalis. Visų pirma, tai viršūninis pumpuras, vaizduojantis ūglio augimo kūgį. IN lapų pažastys sėkliniuose augaluose susidaro pažastiniai ar šoniniai pumpurai. Jei jie išsivysto vienas virš kito (sausmedis, Graikinis riešutas, Robinia ir kt.) Vadinami serijiniais. Jei pumpurai vystosi lapų pažastyse vienas šalia kito (slyvos, javai ir kt.), Tada jie vadinami užstatu. Inkstai gali susiformuoti vidaus organų srityje. Šie inkstai vadinami atsitiktiniais inkstais.
Šalto ir vidutinio klimato medžiuose ir krūmuose susidaro žiemojantys arba neveiklūs pumpurai, kurie dažnai vadinami akimis. Kitais metais iš šių pumpurų išsivysto nauji ūgliai. Išoriniai šių pumpurų lapai dažniausiai išsivysto į inkstų žvynelius, apsaugančius vidines pumpuro dalis nuo pažeidimų.
Žiemos miego, arba snaudžiantys pumpurai taip pat susidaro daugiamečių žolių, ant tų organų, kurie neišnyksta žiemai, t.y. ant šakniastiebių, prie stiebų pagrindo ir kt. Šie pumpurai vadinami atsinaujinimo pumpurais. Pavasarį iš jų vystosi oro ūgliai.
Visi aukščiau išvardyti pumpurai vadinami vegetatyviniais. Tokie inkstai susideda iš viršūnės, rudimentinių mazgų, pradinių tarpuplaukių, lapų primordija, virš kurių gali išsivystyti inkstų primordija, ir rudimentiniai lapai.
Iš inksto, kuriame nėra inkstų primordijos, paprasto ar šakoto Pabėgimas... Iš inksto su inkstų primordija, šakotas Pabėgimas.
Be to, sėkliniai augalai taip pat turi generatyvinius pumpurus. Tai žiedpumpuriai ir pumpurai, iš kurių atsiranda gimnazijos spurgų. Jie skiriasi nuo vegetatyvinės išvaizdos. Be viršūnės, užuomazgų tarpuplaučių ir rudimentinių mazgų, tokie pumpurai turi primordijas, dėl kurių atsiranda gėlių dalys arba kūgių dalys. Prie pumpurų, iš kurių atsiranda žiedynai, susidaro žydinčios primordijos.
Galiausiai yra vadinamieji mišrūs pumpurai, iš kurių susidaro lapiniai ūgliai su žiedais.
Morfologinės savybės pabėgimas reiškia mazgų, tarpinių mazgų, pumpurų struktūros aprašymą. Turi būti nurodytas lapų išdėstymo tipas. Daugumoje augalų jis yra pakaitinis - mazge yra vienas lapas, tačiau jis gali būti priešingas arba susuktas. Tam tikro tipo lapų išdėstymas sudaro lapų mozaiką, o tai leidžia geriausias būdas naudokite erdvę, kad būtų užtikrintas vienodas lapo apšvietimas.
Lapų suskirstymas į tris kategorijas taip pat susijęs su ūglio augimo ir vystymosi procesu: apatiniai lapai, viduriniai lapai, viršūniniai arba viršutiniai lapai. Morfologiniame lapų aprašyme paprastai aprašomi viduriniai lapai, tačiau norint visiškai apibūdinti morfologiją, reikia atskirai aprašyti visų kategorijų lapus, nes net viduriniai lapai skirtumai vieno šūvio metu. Šis reiškinys vadinamas heterofilija arba margumu.
Viršūninis ūglių augimas - ūglio ilgio augimas dėl augimo kūgio modifikavimo, pradinių lapų pradžia ir augimas jo pagrinde. Modifikacijos metu augimo kūgis ilgėja, tampa sudėtingesnis ir keičia savo formą.
Bud... Tai elementarus pabėgimas. Jį sudaro meristematinė ašis, besibaigianti augimo kūgiu (rudimentinis stiebas), ir lapų primordija (rudimentiniai lapai), tai yra, iš pradinių metamerų serijos. Žemiau esantys diferencijuoti lapai dengia augimo kūgį ir primordiją. Taip veikia vegetatyvinis inkstas. Vegetatyviniame-reprodukciniame pumpure augimo kūgis buvo paverstas rudimentine gėle arba rudimentiniu žiedynu. Reprodukciniai (žiediniai) pumpurai susideda tik iš rudimentinės gėlės arba žiedyno ir neturi fotosintezės lapų užuomazgų.
13. Metamorfozuoti ūgliai.
Jų išvaizda dažnai siejama su atsarginių produktų konteinerio funkcijų atlikimu, nepalankių metų sąlygų perkėlimu ir vegetatyviniu dauginimu.
Šakniastiebis- Tai daugiametis požeminis šūvis su horizontalia, kylančia ar vertikalia augimo kryptimi, atliekantis atsarginių produktų kaupimo, atsinaujinimo, vegetatyvinio dauginimosi funkcijas. Šakniastiebyje yra sumažėję lapai žvynelių, pumpurų ir atsitiktinių šaknų pavidalu. Stiebo dalyje kaupiasi atsarginiai produktai. Augimas ir šakojimas vyksta taip pat, kaip ir įprasto ūglio metu. Šakniastiebis nuo šaknies skiriasi tuo, kad yra lapų ir nėra šaknies dangtelio viršūnėje. Šakniastiebis gali būti ilgas ir plonas (kviečių žolė) arba trumpas ir storas. Kasmetiniai oriniai ūgliai susidaro iš viršūninių ir pažastinių pumpurų. Senos šakniastiebio dalys palaipsniui nyksta. Augalai su horizontaliais ilgais šakniastiebiais, sudarančiais daug oro ūglių, greitai užima didelį plotą, o jei tai piktžolės (kviečių žolės), jas kontroliuoti gana sunku. Tokie augalai naudojami smėliui tvirtinti (smaigalys, aristida). Pievose javai, turintys ilgus horizontalius šakniastiebius, vadinami šakniastiebiais (sulenkta žolė, mėlynažolė), o su trumpais - krūminiais (motiejus, baltasis). Šakniastiebiai daugiausia randami daugiamečiuose žoliniuose augaluose, bet kartais krūmuose (euonymus) ir krūmuose (bruknėse, mėlynėse).
Gumbas- Tai sustorėjusi šaudymo dalis, rezervinių produktų talpykla. Gumbai yra antžeminiai ir požeminiai.
Oro gumbas yra pagrindinio (kopūstų) ar šoninio (atogrąžų orchidėjų) ūglio sustorėjimas ir turi normalius lapus.
Požeminis gumbas- hipokotilo (ciklamenų) ar trumpalaikio požeminio ūglio - stolono (bulvės) sustorėjimas. Lapai ant požeminio gumbų sumažėja, jų pažastyse yra pumpurų, vadinamų akimis.
Antžeminis stolonas- Tai trumpalaikis šliaužiantis ūglis, skirtas paskirstymui (teritorijos užgrobimui) ir vegetatyviniam dauginimuisi. Jis turi ilgus tarplapius ir žalius lapus. Ant mazgų susidaro atsitiktinės šaknys, o iš viršūninio pumpuro susidaro sutrumpintas ūglis (rozetė), kuris, mirus stolonui, toliau egzistuoja savarankiškai. Antžeminis stolonas auga simpatijoms. Antžeminiai stolonai, praradę fotosintezės funkciją ir atliekantys daugiausia vegetatyvinio dauginimosi funkciją, kartais vadinami ūsais (braškėmis).
Lemputė- Tai sutrumpintas stiebas (apačia), turintis daugybę, arti esančių lapų ir atsitiktinių šaknų. Dugno viršuje yra inkstas. Daugelyje augalų (svogūnų, tulpių, hiacintų ir kt.) Iš šio pumpuro susidaro šaudymas iš oro, o iš šoninio pažasties pumpuro susidaro nauja lemputė. Išorinės svarstyklės dažniausiai būna sausos, plėvelinės ir atlieka apsauginę funkciją, vidinės - mėsingos, pripildytos atsarginių produktų. Lemputės forma yra sferinė, kiaušinio formos, suplota ir kt.
Corm išoriškai panašus į svogūną, tačiau visos jo lapų žvyneliai yra sausi, o atsarginiai produktai nusėda stiebo dalyje (šafranas, kardelis).
Erškėčiai turėti skirtingą kilmę - nuo ūglio (obuolio, kriaušės, juodadario, gudobelės, gledicijos, citrusinių vaisių), lapo (raugerškio) arba jo dalių: rachis (astragalus), šapalo (baltos akacijos), plokštelės dalies (Compositae). Stuburėliai būdingi augalams karštose, sausose buveinėse.
Antenos susidaro iš ūglio (vynuogių), lapo ar jo dalių: rachio ir kelių lapų (žirnių), lėkštės (rangas), stipules (sarsaparilla). Jie naudojami pritvirtinti prie atramos.
Filokladai- Tai plokšti, lapiniai ūgliai, išsidėstę sumažėjusių lapų pažastyse. Ant jų susidaro gėlės. Augaluose jie randami daugiausia sausose buveinėse (mėsininkas, fillantas). Žvejybos aparatai- vabzdžiaėdžiams augalams būdingi modifikuoti lapai (saulėgrąžos, musmirės). Jie yra ąsočių, urnų, burbuliukų arba užspaudžiamų ir ridenamų plokščių pavidalo. Maži vabzdžiai, patekę į juos, miršta, ištirpsta fermentų pagalba ir augalai juos sunaudoja kaip papildomą mineralų šaltinį.
Šaknis yra požeminis ašinis augalų elementas, kuris yra svarbiausia jų dalis, pagrindinis vegetatyvinis organas. Šaknies dėka augalas įsitvirtina dirvožemyje ir laikomas jame per visą gyvavimo ciklą, taip pat aprūpinamas jame esančiu vandeniu, mineralais ir maistinėmis medžiagomis. Yra įvairių rūšių ir rūšių šaknys. Kiekvienas iš jų turi savo išskirtines savybes. Šiame straipsnyje apžvelgsime esamus šaknų tipus, šaknų sistemų tipus. Taip pat susipažinsime su jiems būdingais bruožais.
Kokios šaknų rūšys egzistuoja?
Standartinė šaknis pasižymi filiformine arba siaura cilindrine forma. Daugelyje augalų, be pagrindinės (pagrindinės) šaknies, vystosi ir kitų rūšių šaknys - šoninės ir atsitiktinės. Pažvelkime atidžiau, kas jie yra.
Pagrindinė šaknis
Šis augalo organas vystosi iš sėklos embriono šaknies. Pagrindinė šaknis visada yra viena (kitos augalų šaknų rūšys dažniausiai yra daugiskaitos). Jis laikomas augale per visą jo gyvavimo ciklą.
Šaknis pasižymi teigiamu geotropizmu, tai yra, dėl gravitacijos gilėja į substratą vertikaliai žemyn.
Adventinės šaknys
Papildomos sąlygos yra augalų šaknų rūšys, susidarančios kituose jų organuose. Šie organai gali būti stiebai, lapai, ūgliai ir tt Pavyzdžiui, javai turi vadinamąsias pirmines atsitiktines šaknis, kurios yra išdėstytos sėklos embriono stiebe. Jie vystosi sėklų daigumo procese beveik vienu metu su pagrindine šaknimi.
Taip pat yra lapinių atsitiktinių šaknų tipų (susidariusių dėl lapų įsišaknijimo), stiebo ar mazgelio (susiformavusių iš šakniastiebių, antžeminių ar požeminių stiebų mazgų) ir kt. Apatiniuose mazguose susidaro galingos šaknys, kurios vadinamos oru ( arba palaikyti) šaknis.
Atsitiktinių šaknų išvaizda lemia augalo gebėjimą daugintis vegetatyviniu būdu.
Šoninės šaknys
Šoninėmis šaknimis vadinamos šaknys, atsirandančios kaip šoninė šaka. Jie gali formuotis tiek pagrindinėse, tiek atsitiktinėse šaknyse. Be to, jie taip pat gali išsišakoti iš šoninių, dėl to susidaro aukštesnių pakopų (pirmosios, antrosios ir trečiosios) šoninės šaknys.
Dideliems šoniniams organams būdingas skersinis geotropizmas, tai yra, jų augimas vyksta beveik horizontalioje padėtyje arba kampu į dirvos paviršių.
Kas vadinama šaknų sistema?
Šaknų sistema vadinama visų rūšių ir tipų šaknys, esančios viename augale (tai yra jų visuma). Priklausomai nuo pagrindinių, šoninių ir atsitiktinių šaknų augimo santykio, nustatomas jo tipas ir pobūdis.
Šaknų sistemų tipai
Jei pagrindinė šaknis yra labai gerai išvystyta ir matoma tarp kitos rūšies šaknų, tai reiškia, kad augalas turi čiaupų sistemą. Tai būdinga daugiausia dviskilčiams augalams.
Šio tipo šaknų sistema išsiskiria giliu daigumu dirvožemyje. Pavyzdžiui, kai kurių žolių šaknys gali prasiskverbti į 10–12 metrų gylį (paršavedžių erškėčiai, liucernos). Medžių šaknų įsiskverbimo gylis kai kuriais atvejais gali siekti 20 m.
Jei atsitiktinės šaknys yra ryškesnės, vystosi daug, o pagrindinei būdingas lėtas augimas, tada susidaro šaknų sistema, vadinama pluoštine.

Paprastai kai kurioms žolinėms augalams būdinga tokia sistema. Nepaisant to, kad pluoštinės sistemos šaknys neprasiskverbia taip giliai, kaip lazdelių sistemos šaknys, jos geriau pina greta esančias dirvožemio daleles. Daugelis purių krūmų ir šakniastiebių žolių, sudarančių gausų pluoštinių plonų šaknų kiekį, yra plačiai naudojami įtvirtinti dauboms, dirvožemiui šlaituose ir tt Geriausios žolės yra laužas be laužo, eraičinas ir kt.

Modifikuotos šaknys
Be aukščiau aprašytų tipiškų, yra ir kitų tipų šaknų ir šaknų sistemų. Jie vadinami modifikuotais.
Saugojimo šaknys
Sandėlyje yra šakniavaisiai ir šakniagumbiai.
Šakniavaisis yra pagrindinės šaknies sustorėjimas dėl maistinių medžiagų nusėdimo. Apatinė stiebo dalis taip pat dalyvauja formuojant šakniavaisį. Susideda daugiausia iš sandėliavimo pagrindo audinio. Šakninių daržovių pavyzdžiai yra petražolės, ridikai, morkos, burokėliai ir kt.

Jei šoninės ir atsitiktinės šaknys yra sustorėjusios laikymo šaknys, tai jos vadinamos šakniagumbiais (kūgiais). Jie vystomi bulvėse, saldžiose bulvėse, jurginuose ir kt.
Oro šaknys
Tai šoninės šaknys, augančios oro dalyje. Yra daugelyje atogrąžų augalų. Iš jo absorbuojamas vanduo ir deguonis oro aplinka... Jie randami atogrąžų augaluose, augančiuose mineralų trūkumo sąlygomis.

Kvėpavimo šaknys
Tai šoninių šaknų rūšis, auganti aukštyn, kylanti virš substrato paviršiaus, vandens. Šio tipo šaknys formuojasi augaluose, augančiuose per drėgnoje dirvoje, pelkėmis. Tokių šaknų pagalba augmenija iš oro gauna trūkstamą deguonį.
Palaikančios (į lentą panašios) šaknys
Šios medžių šaknų rūšys būdingos didelėms rūšims (buko, guobos, tuopos, atogrąžų ir kt.). Jie yra trikampiai vertikalūs ataugos, suformuotos iš šoninių šaknų ir besitęsiančios šalia arba virš dirvos paviršiaus. Jie taip pat vadinami lentomis, nes jie primena lentas, kurios atsiremia į medį.
Siurbimo šaknys (haustoria)
Tai yra papildomų atsitiktinių šaknų rūšis, atsirandanti ant vijoklinių augalų stiebo. Su jų pagalba augalai turi galimybę prisitvirtinti prie tam tikros atramos ir lipti (pinti) aukštyn. Tokios šaknys randamos, pavyzdžiui, atkakliame fikuse, gebenėje ir kt.

Ištraukiamos (susitraukiančios) šaknys
Būdinga augalams, kurių šaknis smarkiai sumažėja išilgine kryptimi prie pagrindo. Pavyzdys yra augalai, turintys svogūnėlių. Ištraukiamos šaknys suteikia svogūnėliams ir šakniavaisiams šiek tiek įdubimų į dirvą. Be to, jų buvimas lemia tvirtą rozetių (pavyzdžiui, kiaulpienių) prigludimą prie žemės, taip pat vertikalios šakniastiebio ir šaknies kaklelio požeminę padėtį.
Mikorizė (grybelio šaknis)
Mikorizė vadinama aukštesnių augalų šaknų simbioze (abipusiai naudingu sugyvenimu) su grybelinėmis hifomis, kurios jas pina, atlikdamos šaknų plaukelių funkcijas. Grybai aprūpina augalus vandeniu ir jame ištirpusiomis maistinėmis medžiagomis. Augalai savo ruožtu aprūpina grybus jų gyvenimui reikalingomis organinėmis medžiagomis.
Mikorizė būdinga daugelio aukštesnių augalų šaknims, ypač sumedėjusiems.
Bakteriniai mazgeliai
Tai modifikuotos šoninės šaknys, pritaikytos simbiotiniam gyvenimui su azotą fiksuojančiomis bakterijomis. Mazgeliai susidaro dėl įsiskverbimo į jaunas šaknis. Šis abipusiai naudingas bendras gyvenimas leidžia augalams gauti azoto, kurį bakterijos perneša iš oro į jiems prieinamą formą. Bakterijoms suteikiama ypatinga buveinė, kurioje jos gali veikti nekonkuruodamos su kitų tipų bakterijomis. Be to, jie naudoja medžiagas, esančias augmenijos šaknyse.

Bakteriniai mazgeliai būdingi ankštinių šeimos augalams, kurie plačiai naudojami kaip meliorantai sėjomainoje, siekiant praturtinti dirvą azotu. Geriausi azotą fiksuojantys augalai yra ankštiniai ankštiniai augalai, tokie kaip mėlyna ir geltona liucernos, raudonieji ir espafonai, ragai ir kt.
Be minėtų metamorfozių, yra ir kitų rūšių šaknų, tokių kaip atraminės šaknys (padedančios sustiprinti stiebą), šakotosios šaknys (padedančios augalams nepaskęsti skystame purve) ir šaknų atžalos (turi atsitiktinių pumpurų ir užtikrina vegetatyvinį dauginimąsi) .









