Корень и корневые системы. Образование боковых корней
Филогенетически корень возник позже стебля и листа - в связи с переходом растений к жизни на суше и вероятно, произошёл от корнеподобных подземных веточек. У корня нет ни листьев, ни в определённом порядке расположенных почек. Для него характерен верхушечный рост в длину, боковые разветвления его возникают из внутренних тканей, точка роста покрыта корневым чехликом. Корневая система формируется на протяжении всей жизни растительного организма. Иногда корень может служить местом отложения в запас питательных веществ. В таком случае он видоизменяется.
Виды корней
Главный корень образуется из зародышевого корешка при прорастании семени. От него отходят боковые корни.
Придаточные корни развиваются на стеблях и листьях.
Боковые корни представляют собой ответвления любых корней.
Каждый корень (главный, боковые, придаточные) обладает способностью к ветвлению, что значительно увеличивает поверхность корневой системы, а это способствует лучшему укреплению растения в почве и улучшению его питания.
Типы корневых систем
Различают два основных типа корневых систем: стержневая, имеющая хорошо развитый главный корень, и мочковатая. Мочковатая корневая система состоит из большого числа придаточных корней, одинаковых по величине. Вся масса корней состоит из боковых или придаточных корешков и имеет вид мочки.
Сильно разветвлённая корневая система образует огромную поглощающую поверхность. Например,
- общая длина корней озимой ржи достигает 600 км;
- длина корневых волосков — 10 000 км;
- общая поверхность корней — 200 м 2 .
Это во много раз превышает площадь надземной массы.

Если у растения хорошо выражен главный корень и развиваются придаточные корни, то формируется корневая система смешанного типа (капуста, помидор).
Внешнее строение корня. Внутреннее строение корня
Зоны корня

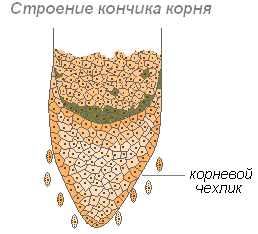
Корневой чехлик
Корень растёт в длину своей верхушкой, где находятся молодые клетки образовательной ткани. Растущая часть покрыта корневым чехликом, защищающим кончик корня от повреждений, и облегчает продвижение корня в почве во время роста. Последняя функция осуществляется благодаря свойству внешних стенок корневого чехлика покрываться слизью, что уменьшает трение между корнем и частичками почвы. Могут даже раздвигать частички почвы. Клетки корневого чехлика живые, часто содержат зёрна крахмала. Клетки чехлика постоянно обновляются за счёт деления. Участвует в положительных геотропических реакциях (направление роста корня к центру Земли).
Клетки зоны деления активно делятся, протяженность этой зоны у разных видов и у разных корней одного и того же растения неодинакова.
За зоной деления расположена зона растяжения (зона роста). Протяжённость этой зоны не превышает нескольких миллиметров.
По мере завершения линейного роста наступает третий этап формирования корня — его дифференциация, образуется зона дифференциации и специализации клеток (или зона корневых волосков и всасывания). В этой зоне уже различают наружный слой эпиблемы (ризодермы) с корневыми волосками, слой первичной коры и центральный цилиндр.
Строение корневого волоска
Корневые волоски — это сильно удлинённые выросты наружных клеток, покрывающих корень. Количество корневых волосков очень велико (на 1 мм 2 от 200 до 300 волосков). Их длина достигает 10 мм. Формируются волоски очень быстро (у молодых сеянцев яблони за 30-40 часов). Корневые волоски недолговечны. Они отмирают через 10-20 дней, а на молодой части корня отрастают новые. Это обеспечивает освоение корнем новых почвенных горизонтов. Корень непрерывно растёт, образуя всё новые и новые участки корневых волосков. Волоски могут не только поглощать готовые растворы веществ, но и способствовать растворению некоторых веществ почвы, а затем всасывать их. Участок корня, где корневые волоски отмерли, некоторое время способен всасывать воду, но затем покрывается пробкой и теряет эту способность.
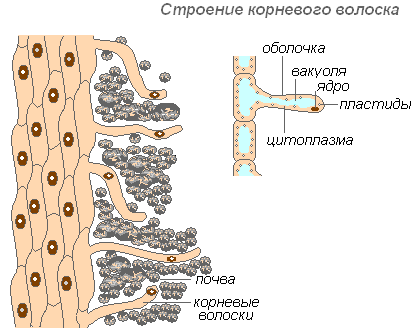
Оболочка волоска очень тонкая, что облегчает поглощение питательных веществ. Почти всю клетку волоска занимает вакуоль, окружённая тонким слоем цитоплазмы. Ядро находится в верхней части клетки. Вокруг клетки образуется слизистый чехол, который содействует склеиванию корневых волосков с частицами почвы, что улучшает их контакт и повышает гидрофильность системы. Поглощению способствует выделение корневыми волосками кислот (угольной, яблочной, лимонной), которые растворяют минеральные соли.
Корневые волоски играют и механическую роль — они служат опорой верхушке корня, которая проходит между частичками почвы.
Под микроскопом на поперечном срезе корня в зоне всасывания видно его строение на клеточном и тканевом уровнях. На поверхности корня — ризодерма, под ней — кора. Наружный слой коры — экзодерма, вовнутрь от неё — основная паренхима. Её тонкостенные живые клетки выполняют запасающую функцию, проводят растворы питательных веществ в радиальном направлении — от всасывающей ткани к сосудам древесины. В них же происходит синтез ряда жизненно важных для растения органических веществ. Внутренний слой коры — эндодерма. Растворы питательных веществ, поступающие из коры в центральный цилиндр через клетки эндодермы, проходят только через протопласт клеток.

Кора окружает центральный цилиндр корня. Она граничит со слоем клеток, долго сохраняющих способность к делению. Это перицикл. Клетки перицикла дают начало боковым корням, придаточным почкам и вторичным образовательным тканям. Вовнутрь от перицикла, в центре корня, находятся проводящие ткани: луб и древесина. Вместе они образуют радиальный проводящий пучок.
Проводящая система корня проводит воду и минеральные вещества из корня в стебель (восходящий ток) и органические вещества из стебля в корень (нисходящий ток). Состоит она из сосудисто-волокнистых пучков. Основными слагаемыми частями пучка являются участки флоэмы (по ним вещества передвигаются к корню) и ксилемы (по которым вещества передвигаются от корня). Основные проводящие элементы флоэмы — ситовидные трубки, ксилемы — трахеи (сосуды) и трахеиды.
Процессы жизнедеятельности корня
Транспорт воды в корне
Всасывание воды корневыми волосками из почвенного питательного раствора и проведение её в радиальном направлении по клеткам первичной коры через пропускные клетки в эндодерме к ксилеме радиального проводящего пучка. Интенсивность поглощения воды корневыми волосками называется сосущей силой (S), она равна разнице между осмотическим (P) и тургорным (T) давлением: S=P-T.
Когда осмотическое давление равно тургорному (P=T), то S=0, вода перестаёт поступать в клетку корневого волоска. Если концентрация веществ почвенного питательного раствора будет выше, чем внутри клетки, то вода будет выходить из клеток и наступит плазмолиз — растения завянут. Такое явление наблюдается в условиях сухости почвы, а также при неумеренном внесении минеральных удобрений. Внутри клеток корня сосущая сила корня возрастает от ризодермы по направлению к центральному цилиндру, поэтому вода движется по градиенту концентрации (т. е. из места с большей её концентрацией в место с меньшей концентрацией) и создаёт корневое давление, которое поднимает столбик воды по сосудам ксилемы, образуя восходящий ток. Это можно обнаружить на весенних безлистных стволах, когда собирают «сок», или на срезанных пнях. Истекание воды из древесины, свежих пней, листьев, называется «плачем» растений. Когда распускаются листья, то они тоже создают сосущую силу и притягивают воду к себе — образуется непрерывный столбик воды в каждом сосуде — капиллярное натяжение. Корневое давление является нижним двигателем водного тока, а сосущая сила листьев — верхним. Подтвердить это можно с помощью несложных опытов.
Всасывание воды корнями
Цель: выяснить основную функцию корня.
Что делаем: растение, выращенное на влажных опилках, отряхнём его корневую систему и опустим в стакан с водой его корни. Поверх воды для защиты её от испарения нальём тонкий слой растительного масла и отметим уровень.

Что наблюдаем: через день-два вода в ёмкости опустилась ниже отметки.

Результат: следовательно, корни всосали воду и подали её наверх к листьям.
Можно ещё проделать один опыт, доказывающий всасывание питательных веществ корнем.
Что делаем: срежем у растения стебель оставив пенёк высотой 2-3 см. На пенёк наденем резиновую трубку длиной 3 см, а на верхний конец наденем изогнутую стеклянную трубку высотой 20-25 см.
Что наблюдаем: вода в стеклянной трубке поднимается, и вытекает наружу.
Результат: это доказывает, что воду из почвы корень всасывает в стебель.

А влияет ли температура воды на интенсивность всасывания корнем воды?
Цель: выяснить, как температура влияет на работу корня.
Что делаем: один стакан должен быть с тёплой водой (+17-18ºС), а другой с холодной (+1-2ºС).
Что наблюдаем: в первом случае вода выделяется обильно, во втором — мало, или совсем приостанавливается.
Результат: это является доказательством того, что температура сильно влияет на работу корня.

Тёплая вода активно поглощается корнями. Корневое давление повышается.

Холодная вода плохо поглощается корнями. В этом случае корневое давление падает.
Минеральное питание
Физиологическая роль минеральных веществ очень велика. Они являются основой для синтеза органических соединений, а также факторами, которые изменяют физическое состояние коллоидов, т.е. непосредственно влияют на обмен веществ и строение протопласта; выполняют функцию катализаторов биохимических реакций; воздействуют на тургор клетки и проницаемость протоплазмы; являются центрами электрических и радиоактивных явлений в растительных организмах.
Установлено, что нормальное развитие растений возможно только при наличии в питательном растворе трёх неметаллов — азота, фосфора и серы и — и четырёх металлов — калия, магния, кальция и железа. Каждый из этих элементов имеет индивидуальное значение и не может быть заменён другим. Это макроэлементы, их концентрация в растении составляет 10 -2 –10%. Для нормального развития растений нужны микроэлементы, концентрация которых в клетке составляет 10 -5 –10 -3 %. Это бор, кобальт, медь, цинк, марганец, молибден др. Все эти элементы есть в почве, но иногда в недостаточном количестве. Поэтому в почву вносят минеральные и органические удобрения.
Растение нормально растёт и развивается в том случае, если в окружающей корни среде будут содержаться все необходимые питательные вещества. Такой средой для большинства растений является почва.
Дыхание корней
Для нормального роста и развития растения необходимо чтобы к корню поступал свежий воздух. Проверим, так ли это?
Цель: нужен ли воздух корню?
Что делаем: возьмём два одинаковых сосуда с водой. В каждый сосуд поместим развивающие проростки. Воду в одном из сосудов каждый день насыщаем воздухом с помощью пульверизатора. На поверхность воды во втором сосуде нальём тонкий слой растительного масла, так как оно задерживает поступление воздуха в воду.

Что наблюдаем: через некоторое время растение во втором сосуде перестанет расти, зачахнет, и в конце концов погибнет.

Результат: гибель растения наступает из-за недостатка воздуха, необходимого для дыхания корня.
Видоизменения корней
У некоторых растений в корнях откладываются запасные питательные вещества. В них накапливаются углеводы, минеральные соли, витамины и другие вещества. Такие корни сильно разрастаются в толщину и приобретают необычный внешний вид. В формировании корнеплодов участвуют и корень, и стебель.
Корнеплоды
Если запасные вещества накапливаются в главном корне и в основании стебля главного побега, образуются корнеплоды (морковь). Растения, образующие корнеплоды, в основном двулетники. В первый год жизни они не цветут и накапливают в корнеплодах много питательных веществ. На второй — они быстро зацветают, используя накопленные питательные вещества и образуют плоды и семена.
Корневые клубни

У георгина запасные вещества накапливаются в придаточных корнях, образуя корневые клубни.
Бактериальные клубеньки
Своеобразно изменены боковые корни у клевера, люпина, люцерны. В молодых боковых корешках поселяются бактерии, что способствует усвоению газообразного азота почвенного воздуха. Такие корни приобретают вид клубеньков. Благодаря этим бактериям эти растения способны жить на бедных азотом почвах и делать их более плодородными.
Ходульные

У пандуса, произрастающего в приливно-отливной зоне, развиваются ходульные корни. Они высоко над водой удерживают на зыбком илистом грунте крупные облиственные побеги.
Воздушные

У тропических растений, живущих на ветвях деревьев, развиваются воздушные корни. Они часто встречаются у орхидей, бромелиевых, у некоторых папоротников. Воздушные корни свободно висят в воздухе, не достигая земли и поглощая попадающую на них влагу от дождя или росы.
Втягивающие

У луковичных и клубнелуковичных растений, например у крокусов, среди многочисленных нитевидных корней имеется несколько более толстых, так называемых втягивающих, корней. Сокращаясь, такие корни втягивают клубнелуковицу глубже в почву.
Столбовидные

У фикуса развиваются столбовидные надземные корни, или корни-подпорки.
Почва как среда обитания корней
Почва для растений является средой, из которой оно получает воду и элементы питания. Количество минеральных веществ в почве зависит от специфических особенностей материнской горной породы, деятельности организмов, от жизнедеятельности самих растений, от типа почвы.
Почвенные частицы конкурируют с корнями за влагу, удерживая её своей поверхностью. Это так называемая связанная вода, которая подразделяется на гигроскопическую и плёночную. Удерживается она силами молекулярного притяжения. Доступная растению влага представлена капиллярной водой, которая сосредоточена в мелких порах почвы.
Между влагой и воздушной фазой почвы складываются антагонистические отношения. Чем больше в почве крупных пор, тем лучше газовый режим этих почв, тем меньше влаги удерживает почва. Наиболее благоприятный водно-воздушный режим поддерживается в структурных почвах, где вода и воздух находятся одновременно и не мешают друг другу — вода заполняет капилляры внутри структурных агрегатов, а воздух — крупные поры между ними.
Характер взаимодействия растения и почвы в значительной степени связан с поглотительной способностью почвы — способностью удерживать или связывать химические соединения.
Микрофлора почвы разлагает органические вещества до более простых соединений, участвует в формировании структуры почвы. Характер этих процессов зависит от типа почвы, химического состава растительных остатков, физиологических свойств микроорганизмов и других факторов. В формировании структуры почвы принимают участие почвенные животные: кольчатые черви, личинки насекомых и др.
В результате совокупности биологических и химических процессов в почве образуется сложный комплекс органических веществ, который объединяют термином «гумус».
Метод водных культур
В каких солях нуждается растение, и какое влияние оказывают они на рост и развитие его, было установлено на опыте с водными культурами. Метод водных культур — это выращивание растений не в почве, а в водном растворе минеральных солей. В зависимости от поставленной цели в опыте можно исключить отдельную соль из раствора, уменьшить или увеличить ее содержание. Было выяснено, что удобрения, содержащие азот, способствуют росту растений, содержащие фосфор — скорейшему созреванию плодов, а содержащие калий — быстрейшему оттоку органических веществ от листьев к корням. В связи с этим содержащие азот удобрения рекомендуется вносить перед посевом или в первой половине лета, содержащие фосфор и калий — во второй половине лета.
С помощью метода водных культур удалось установить не только потребность растения в макроэлементах, но и выяснить роль различных микроэлементов.
В настоящее время известны случаи, когда выращивают растения методами гидропоники и аэропоники.
Гидропоника — выращивание растений в сосудах, заполненных гравием. Питательный раствор, содержащий необходимые элементы, подаётся в сосуды снизу.
Аэропоника — это воздушная культура растений. При этом способе корневая система находится в воздухе и автоматически (несколько раз в течение часа) опрыскивается слабым раствором питательных солей.
ОРГАНОГРАФИЯ
Органография – описание органов живого организма.
Орган – часть организма, дифференцированная морфологически и специализированная функционально. В соответствии с двумя главными функциями: поддержание жизни данного растения и воспроизведение его в ряде последующих поколений – выделяют вегетативные и генеративные (репродуктивные) органы.
Вначале мы будем изучать вегетативные органы, образующие систему побега и корневую систему. Элементом корневой системы является корень.
Корень
Корень эволюционно возник позже побега и происходит от ризоидов первых наземных растений – риниофитов.
Разные части корня выполняют неодинаковые функции и характеризуются определенными морфологическими особенностями. Эти части получили название зон. Зоны молодого корня: зона деления, зона роста или растяжения, зона поглощения или всасывания и зона проведения.
Снаружи апикальная меристема прикрыта корневым чехликом. Живые паренхимные клетки чехлика продуцируют слизь. Слизистое вещество представляет собой сильно гидратированный полисахарид пектиновой природы. Он накапливается в пузырьках диктиосом, которые, сливаясь с плазмолеммой, высвобождают его в пространство между плазмолеммой и клеточной оболочкой. Затем слизь выходит на поверхность клеточной оболочки, где и образует мелкие капли.
Благодаря такому слизистому покрытию частички почвы прилипают к кончику корня и корневым волоскам. Слизь служит в качестве абсорбирующей поверхности, которая осуществляет обмен ионов. Слизистое покрытие охраняет кончик корня от вредных веществ почвы и от высыхания, создает благоприятные условия для поселения полезных бактерий.
По мере роста корня и проталкивания корневого чехлика вперед клетки на периферии чехлика слущиваются. Продолжительность жизни клеток от возникновения до слущивания составляет 4-9 дней в зависимости от длины чехлика и вида растения.
Корневой чехлик контролирует георецепцию корня. Место восприятия гравитации в чехлике – центральная колонка клеток – колумелла, в которой амилопласты действуют как статолиты (рецепторы гравитации).
Апикальная меристема состоит из мелких (10 – 20 мкм в диаметре) многогранных клеток с густой цитоплазмой и крупными ядрами.
У споровых растений моноинициальный тип апекса корня.
В корнях семенных растений описаны два основных типа апикальной организации. В одном из них все области имеют общие инициали. Это открытый тип апикальной меристемы, который, вероятно, является филогенетически примитивным. Во втором типе каждая из трех частей, т.е. центральный (или проводящий) цилиндр, первичная кора и корневой чехлик, имеют свои собственные инициали. Это закрытый тип апикальной меристемы.
Инициали, ответственные за организацию клеточного роста на первых стадиях, в основном утрачивают митотическую активность на последующих стадиях роста корня. Вместо них митотическую активность проявляют клетки, расположенные несколько глубже в теле корня. Эти наблюдения послужили поводом для формирования теории покоящегося центра в апикальной меристеме. Согласно этой теории, самые дистальные клетки (дистальный – наиболее удаленная от центральной точки часть органа: нижний конец корня) в теле корня (первые инициали плеромы и периблемы) делятся редко, имеют примерно одинаковые размеры и низкие скорости синтеза НК и белка. Покоящийся центр имеет полусферическую или дисковидную форму, варьирует в объеме в зависимости от размера корня. Клетки покоящегося центра делятся приблизительно в 10 раз медленнее, чем соседние клетки.
В корнях, эспериментально поврежденных, покоящийся центр способен восстанавливать меристему. Т.е., покоящийся центр играет существенную роль в формировании структуры и в развитии корня.
Особенностью кончика корня является размещение продольных рядов клеток, берущих начало в апикальной меристеме.
Слои апикальной меристемы корня:
Дерматоген (греч. derma – кожица, genos – рождение) – наружный слой верхушечной меристемы, дифференцируется в ризодерму. Клетки дерматогена правильной формы.
Плерома (греч. pleroma – заполнение) – центральная часть верхушечной меристемы, дифференцируется в клетки центрального цилиндра. Клетки вытянуты по оси.
Периблема (греч. periblema – покров) – слой верхушечной меристемы между дерматогеном и плеромой, дифференцируется в клетки первичной коры.
Первичная кора дифференцируется на уровне зоны растяжения. В зоне полгощения на долю первичной коры приходится большая часть поперечного сечения. Называется "первичной" благодаря происхождению из первичной меристемы, "корой" – благодаря периферическому расположению.
Наружные клетки первичной коры, расположенные под ризодермой, называются экзодермой. Внутренний слой первичной коры – эндодерма.
Основная масса первичной коры – мезодерма. Мезодерма образована паренхимными клетками, расположенными рыхло, что способствует аэрации клеток корня.
Корни голосеменных и большинства двудольных рано сбрасывают первичную кору. У них клетки первичной коры остаются паренхиматозными. В противоположность этому первичная кора однодольных сохраняется на протяжении всей жизни корня, и ее клетки образуют вторичные лигнифицированные оболочки.
Центр корня занимает осевой цилиндр – стела. Начинает дифференцироваться в зоне роста. Формирование осевого цилиндра начинается с образования наружного его слоя - перицикла. Перицикл относится к первичным латеральным меристемам, клетки его долго сохраняют меристематическую активность. Обычно перицикл представлен одним слоем клеток, хотя бывает и многослойным.
У большинства семенных растений в перицикле закладываются боковые корни, возникают зачатки придаточных почек. У видов со вторичным ростом перицикл участвует в формировании камбия и дает начало первому слою феллогена.
Под перициклом закладываются клетки прокамбия, которые дают начало первичной флоэме, позднее – первичной ксилеме. Количество лучей ксилемы варьирует у разных видов от двух, трех, четырех до нескольких. Соответственно количеству лучей ксилемы корни называют диархными, триархными, тетрархными, полиархными. Первичными считаются тетрархные корни. У однодольных – полиархная стела.
Таким образом, общий план первичного строения корня и морфологическая дифференциация тканей показывают высокую ступень специализации и приспособления корня к функциям всасывания и проведения воды и минеральных веществ.
Образование боковых корней
Боковые корни возникают на периферии центрального цилиндра на разных расстояниях от апикальной меристемы. Т.к. закладываются они в глубоко расположенных тканях корня, их происхождение называют эндогенным (с греч. "рождающийся внутри"). Боковые корни голосеменных и покрытосеменных закладываются в перицикле. Клетки перицикла, участвующие в образовании боковых корней, - корнеродная дуга.
Несколько слоев клеток корневого примордия может образовать и эндодерма.
В диархном корне боковой корень закладывается между флоэмой и ксилемой, в триархном, тетрархном – напротив ксилемы, в полиархном корне однодольных – напротив флоэмы. Клетки перицикла делятся в различных направлениях. В результате чего формируется небольшой корневой бугорок. Увеличиваясь в размерах, зачаток корня давит на эндодерму. Но оболочки эндодермы твердые. Поэтому эндодерма выпячивается. Выпуклость эндодермы называется корневым кармашком.
Примордий корня проходит через первичную кору, секретируя при этом ферменты, разрушающие клетки коры на своем пути. На ранних стадиях развития этот зачаток формирует корневой чехлик, апикальную и первичные меристемы. Со временем за счет дифференциации паренхимных клеток в элементы ксилемы и флоэмы центральные цилиндры бокового и родительского корня соединяются между собой.
У однодольных первичная структура корня сохраняется на протяжении всей жизни и вторичные ткани не возникают. С возрастом могут увеличиваться размеры клеток, утолщаться клеточные оболочки, но общий "план" строения корня остается неизменным.
У голосеменных и двудольных происходят вторичные изменения, и формируется вторичная структура корня. Образование вторичной структуры корня связано прежде всего с деятельностью камбия. Камбий образуется путем деления тех прокамбиальных клеток, которые остаются недифференцированными между первичной флоэмой и первичной ксилемой. Затем клетки перицикла, локализованные снаружи от выступов ксилемы, также проявляют камбиальную активность.
Камбий откладывает к центру клетки вторичной ксилемы, к периферии – клетки вторичной флоэмы. Так образуются проводящие пучки. Их образуется столько, сколько было лучей первичной ксилемы. Под давлением нарастающих вторичных тканей клетки первичной флоэмы расплющиваются. Первичная ксилема сохраняет центральное положение и видна в виде звездочки. Со временем облитерируется и первичная ксилема.
При вторичном строении корня первичные ткани не функционируют как проводящие элементы. Ток веществ в обоих направлениях осуществляется по вторичным тканям.
Камбий, который берет начало в перицикле, образует широкие паренхимные лучи, которые размещаются между открытых коллатеральных пучков. Это первичные сердцевинные лучи. Они обеспечивают физиологическую связь центральной части корня с первичной корой. Позднее могут закладываться более узкие вторичные сердцевинные лучи, соединяющие вторичную ксилему и флоэму.
Вторичные изменения происходят не только в центральном цилиндре. Из-за нарастания в толщину проводящих тканей первичная кора отодвигается наружу, разрывается и сбрасывается вместе с эндодермой.
В наружной части перицикла берет начало феллоген и откладывает наружу феллему, а внутрь – многослойную феллодерму. Таким образом, корни двудольных, имеющих вторичное строение, покрыты перидермой.
В многолетних корнях феллоген возникает в глубине корня. Тогда корень, как и стебель, образует ритидом.
Важный экологический аспект вторичного роста корня представляет собой способность корней различных растений одного и того же вида к естественному срастанию. Там, где корни приходят в контакт один с другим, они объединяются благодаря вторичному росту. При срастании корней устанавливается непрерывность проводящей системы между обоими сросшимися партнерами. Благодаря срастанию корней многие деревья оказываются взаимосвязанными, и если некоторые из них вырубаются, то пни еще долго остаются живыми. Срастание корней способствует быстрой передаче инфекции от одного дерева к другому.
Дифференциация корней в корневых системах проявляется в разных формах. Наибольшее значение имеет дифференциация на главные, боковые и придаточные корни.
В особую группу выделяют корни, возникающие в апикальных меристемах высших споровых растений. Предполагают, что они являются самой архаичной группой корней и сохранили наибольшее сходство с первичными корнями, которые входили в состав единой системы ветвления надземных и подземных осей. Корни, входящие в эту группу, называют апицигенными.
Основываясь на последовательности возникновения корневых систем в процессе эволюции и принадлежности к крупным систематическим группам, можно наметить три эволюционных уровня корневых систем.
I. Корневые системы высших споровых представлены исключительно системами апицигенных корней, последовательно сменяющимися друг друга, что и является основным механизмом обновления в корневых системах. Ветвление верхушечное или боковое, причем не исключено, что оба типа ветвления возникли независимо друг от друга. Нет метаморфозов и корней с узкой специализацией.
П. Корневая система голосеменных отличается наличием главного корня, что можно связать с появлением семени и биполярного зародыша с зародышевым корешком. В механизм обновления включились не только боковые корни подчиненных порядков, но и ризогенные придаточные корни (т.е. придаточные корни на корнях, придаточные корни на побегах не получили у голосеменных заметного развития). Специализированные и метаморфизированные корни единичны.
Ш. Корневая система покрытосеменных характеризуется сильным развитием придаточных корней, а также метаморфозов и корней с узкой специализацией. Механизмы обновления разнообразны и эффективны.
Метаморфозы корневых систем
Метаморфозы корневых систем – это совокупность анатомических и морфологических признаков, выработанных филогенетически и наследственно закрепленных и вызванных изменением физиологических функций. Метаморфоз (metamorphosis, греч. – превращение) – результат изменений, которые происходят на генетическом уровне и которые приводят к морфологическим и функциональным перестройкам органа, системы органов или организма в целом.
I. Метаморфозы, связанные с выполнением нетипичных функций:
1) дыхательные корни;
2) воздушные корни эпифитов;
3) ходульные корни;
4) корни подпорки.
П. Метаморфозы, связанные с гиперфункционированием:
1) корнеплоды;
2) корневые шишки;
3) корневые клубеньки;
4) микориза.
Корнеплоды характерны для двулетних двудольных растений. Это условное название. Развиваются из вегетативных органов и никакого отношения к плодам не имеют. В образовании корнеплода принимают участие нижняя часть стебля и корень.
В первый год жизни растение вегетирует, образуется корнеплод. На второй год формируются цветки, плоды, семена. Значит, корнеплод является органом запаса веществ, что позволяет растению перенести период покоя и завершить развитие на следующий год.
Типы корнеплодов:
1.Монокамбиальный.
Исходным является корень с диархным осевым цилиндром. Образование корнеплода начинается со вторичных изменений, связанных с заложением и работой одного камбиального кольца и дальнейшей паренхиматизацией.
Мы знаем, что камбий во внутрь откладывает вторичную ксилему, наружу – вторичную флоэму. Анатомические отличия касаются места развития запасающей ткани. Если сильное развитие паренхимы происходит во флоэме, образуется флоэмный тип корнеплода – тип морковь. Если в ксилеме – ксилемный тип – тип репа.
2.Поликамбиальный тип корнеплодов.
Закладывается одно типичное камбиальное кольцо, остальные – дополнительные перициклического происхождения. Их может быть до 25. Эти дополнительные кольца камбия образуют открытые коллатеральные проводящие пучки. Чем дальше кольцо от первого камбиального кольца, тем больше запасающей паренхимы оно продуцирует. Пример поликамбиального корнеплода – свекла.
В моно – и поликамбиальных корнеплодах первичная кора рано отмирает и слущивается. Покровной тканью служит перидерма.
Таким образом, общим признаком всех корнеплодов является наличие большого количества запасающей паренхимы, пронизанной проводящими тканями. Тесная связь между проводящими тканями и запасающими достигается благодаря разнообразным модификациям вторичного роста.
У некоторых растений запасающие корни формируются из боковых и придаточных корней в виде корневых шишек. Корневые шишки однодольных (любка) бескамбиальные, имеют первичное строение; двудольных - камбиальные, имеют вторичное строение. Пример: корневые шишки георгина флоэмного типа.
Корневые шишки по всей поверхности образуют придаточные корни, а в верхней части – придаточные почки и поэтому являются органами вегетативного размножения.
Известно, что рост растений чаще всего лимитирует недостаточное обеспечение их фиксированным азотом. Азот в форме стабильной молекулы N 2 составляет 80% атмосферы. Перед фиксацией (восстановлением N 2 до аммиака NH 3) эта молекула должна расщепиться. Фиксация азота осуществляется некоторыми свободноживущими бактериями, которые употребляют органическое вещество почвы. Некоторые бактерии поселяются на корнях высших растений. Растение "привлекает" бактерии при помощи органических корневых выделений. А микроорганизмы в свою очередь обеспечивают растение фиксированным азотом. Такая взаимополезная ассоциация двух организмов называется симбиозом. Чаще всего корни образуют ассоциации с азотфиксирующими бактериями рода Rhizobium. Более 60 семейств двудольных и однодольных растений способны вступать в симбиоз с бактериями. Каждый вид растения имеет собственный симбиотический Rhizobium. Взаимное узнавание растения и бактерии осуществляется путем прикрепления особого белка – лектина, находящегося на поверхности трихобластов, к специфической бактерии. Корневые волоски под влиянием выделяемых бактериями ростовых гормонов деформируются. Бактерии проникают внутрь этих искривленных клеток. Внутри клеток бактерии размножаются, образуя инфекционную нить, заключенную в футляр из камеди. Эта нить проникает глубоко в корень. Результатом такого проникновения бактерий является сильное разрастание клеток мезодермы корня, что приводит к образованию клубеньков. Самая наружная часть клубенька долго остается меристематической зоной, а бактерии размещаются во внутренней части клубенька. Сама бактериальная ткань представлена мелкими клетками с вязкой цитоплазмой и огромным количеством бактерий. Бактериальная ткань окружена проводящими пучками, которые связаны с центральным цилиндром корня. Т.е. существует система транспорта для обмена питательными веществами между бактериями и растениями. Азотфиксирующие бактерии оказывают благоприятное воздействие и на азотное питание растений, и на почвенное плодородие.
Физиологическая активность корня, связанная с обеспечением растения водой и минеральными веществами, может усиливаться путем его симбиотической ассоциации со специфическим грибом. Такой тесный взаимополезный симбиоз корней и грибов и есть микориза. Микориза известна у большинства сосудистых растений. Всего несколько семейств цветковых не образуют ее или образуют редко, например, капустные, осоковые.
Микоризные грибы играют ключевую роль в минеральном питании сосудистых растений. Значительная роль микоризы в поглощении и переносе фосфора.
При двустороннем транспорте (перемещении фосфатов от грибов к растениям и углеводов в обратном направлении) потеря и поглощение происходят в апопластической зоне между плазматическими мембранами растения и гриба.
Микоризные грибы используют те источники Р, которые обычно не доступны растениям. Микориза способствует также поглощению растениями Zn, Mn, Cu. Эти элементы в почве слабоподвижны, и поэтому вокруг корневых волосков быстро образуются обедненные ими зоны. Сеть гифов микоризных грибов распространяется на несколько сантиметров от каждого корня и тем самым увеличивает поверхность поглощения корней.
Различают следующие типы микоризы:
1)эктомикоризы: гриб покрывает весь кончик корня густым чехлом и проникает в межклетники;
2)эндомикоризы: гриб внедряется внутрь клеток.
Эктомикориза характерна для определенных семейств деревьев и кустарников (сосновые, ивовые, буковые). Формируется эктомикориза на молодых, еще не опробковевших корнях. Эктомикоризные корни короткие, разветвленные и выглядят разбухшими.
У таких корней развитие корневых волосков подавлено и объем апикальной меристемы и корневого чехлика может быть уменьшен. Функцию корневых волосков выполняют гифы.
Эндомикоризные корни по форме похожи с обычными корнями, но отличаются от них более темной окраской. Грибные гифы проникают в клетки коры корня, где образуют везикулы (вздутия) и арбускулы (древовидные разветвления). Поэтому эндомикоризу часто называют везикулярно-арбускулярной. В определенных условиях гифы гриба могут "перевариваться" клетками. Корневые волоски на корнях сохраняются. Это наиболее распространенный тип микоризы. Встречается приблизительно у 80% всех сосудистых растений, а именно у травянистых растений, включая зерновые; у кустарников, деревьев (яблони, клены…). У молодого растения тополя, например, формируется везикулярно-арбускулярная эндомикориза, а по мере старения – чехольчатая эктомикориза.
Для вересковых Ericaceae характерна эрикоидно-арбутоидная микориза. Гриб вокруг корня образует мощное скопление гифов, а тонкие боковые гифы проникают в клетки коры корня. Эти микоризные грибы значительно снижают токсичность почв для вересковых, что позволяет этим растениям поселяться на кислых, бедных питательными веществами почвах.
Встречается еще один тип – эндомикориза орхидных. Семена орхидных в природе прорастают только при наличии соответствующих грибов.
Вообще в природных условиях микоризные ассоциации скорее являются правилом, чем исключением.
Исследование остатков древнейших сосудистых растений показало, что эндомикориза встречалась у них не реже, чем у современных растений. Это дало возможность допустить, что возникновение микоризы в процессе эволюции могло быть одним из факторов, позволившим растениям заселить сушу. Учитывая слабое развитие почвы во время этого заселения, можно допустить, что микоризные грибы были основой минерального питания растений. Современные растения-пионеры с микоризой имеют больше шансов на выживание на бедных почвах.
Растения с микотрофным типом питания более устойчивы к наличию тяжелых металлов в окружающей техногенной среде.
Несколько слов о корнях растений влажного тропического леса. Более распространены, чем ходульные, досковидные опорные корни. Корни напоминают доски, прислоненные верхними концами к дереву. В то время как ходульные корни являются придаточными корнями, которые возникают на стволе и растут вниз, образование досковидных корней связано с боковыми корнями, растущими непосредственно под поверхностью почвы. Боковые корни развивают вертикальные выросты, треугольные в поперечном сечении, которые прилегают к стволу дерева.
Древесина досковидных корней часто тверже, чем остальная древесина того же дерева, и нередко они бывают покрыты тонким слоем коры. Судя по кольцам прироста, видным на поперечном разрезе этих корней, вначале это нормальные, округлые в сечении корни, но затем они развивают очень сильное утолщение на верхней стороне. Количество досковидных корней у дерева варьирует от 1 до 10, но в большинстве случаев их три или более. В тропиках образование досковидных корней особенно заметно у деревьев, растущих в болотистых лесах и на плохо дренированных почвах. Нередко досковидные корни используют для промышленных целей. Так, досковидные корни произрастающего в Индонезии дерева Koompassia excelsa идут на изготовление обеденных столов.
ПОБЕГ
Побег, как и корень, является основным вегетативным органом растения. Побег выполняет функцию воздушного питания, может выполнять и ряд других функций и способен к метаморфозам.
Вегетативный невидоизмененный побег состоит из осевой части – стебля; листьев и почек. Главная внешняя черта, отличающая побег от корня, - это олиственность побега.
Участок стебля, который связан с возникновением боковых органов (ветвей, листьев и пр.) – узел. Участок стебля между соседними узлами – междоузлия. Внутренний угол между листом и стеблем – пазуха листа. Каждый повторяющийся элемент побега (узел с листом и почкой в пазухе листа и междоузлие) образует метамер. Побег, таким образом, состоит из серии метамеров, т.е. имеет метамерное строение.
В начале 90-х годов XX века американскими учеными вместо понятий узла и междоузлия для анализа роста и дифференциации растения было предложено понятие – стеблевой элемент. Стеблевой элемент – это участок стебля между краями листовых примордиев. Выявляется на апексе побега, где еще нет междоузлия. Т.е. новое понятие "стеблевой элемент" соответствует понятию "метамер".
Строение апикальной меристемы побега сложней, чем в корне. Ведь в побеге апикальная меристема не только образует клетки первичных тканей, но и участвует в формировании примордиев листьев и пазушных почек, развивающихся затем в боковые побеги. Эта меристема не защищена структурой, подобной корневому чехлику.
Типы апексов такие же, как и в корне: моноинициальный, зональный, ярусный. Количество ярусов больше, чем в корне.
Эволюционно примитивный моноинициальный апекс. Пример такого типа апекса у хвощей. Одиночная крупная вытянутая апикальная клетка сильно вакуолизирована. Делится в направлениях, параллельных своим поверхностям (за исключением самой наружной поверхности). Ближайшие производные клетки также сильно вакуолизированы, но по мере их деления возникают мелкие клетки с густыми протопластами. Такие клетки обнаружены по периферии верхушки стебля, где закладываются листовые зачатки.
Вегетативный апекс побега большинства цветковых построен по типу туники – корпуса – двух зон, отличающихся плоскостями клеточных делений. Туника (лат. - оболочка) включает наружные слои клеток, делящихся антиклинально (перпендикулярно к поверхности) и обеспечивающих увеличение поверхности. Корпус (лат. – тело) – центральная группа клеток под туникой, делятся в разных плоскостях, образуя основной объем побега. Корпус и каждый слой туники имеют свои собственные инициали. В тунике инициали размещаются в центре по оси. Двудольные имеют в основном двуслойную тунику, однодольные – однослойную, реже двуслойную.
В то время как концепция туники – корпуса способствует нашему пониманию расположения и роста клеток в самой апикальной меристеме, распознавание различных по характеру зон клеток в прилегающих к ней областях помогает выявить вклад меристемы в дифференциацию тканей и органов в побеге. Наличие в апикальной меристеме зон, имеющих различные цитологические характеристики, есть цитогистологическая зональность.
Корпус соответствует области клеток, называемой зоной центральных материнских клеток. Клетки этой зоны сильно вакуолизированы – черта, связанная с относительно низкой скоростью митотической активности. Зона центральных материнских клеток окружена периферической меристемой, возникающей частично из туники, частично из корпуса. С наружного слоя туники берет начало гистоген протодерма. Периферическая меристема происходит из латеральных производных апикальных инициалей и из центральных материнских клеток. Сердцевинная меристема формируется в результате делений по периферии центральных материнских клеток в слое, называемом переходной зоной. Клетки периферической зоны имеют густые протопласты, следовательно, обладают высокой митотической активностью.
В определенных местах с особенно высокой активностью закладываются листовые бугорки (или первичные бугорки).
Возникновение нового листового примордия связано с изменением направления делений клеток, которому предшествует изменение ориентации микротрубочек в периферических слоях цитоплазмы. Осмотическое давление также влияет на рост примордия.
В период активного роста апикальная меристема побега образует листовые примордии так быстро, что узлы и междоузлия вначале не отличаются. Постепенно участки между ярусами листьев начинают расти, вытягиваться и приобретают вид междоузлий. В то же время области прикрепления листьев оформляются в узлы.
Ниже, где появляются зачатки листьев, наблюдается вакуолизация. Это указывает на начало дифференциации первичной коры и сердцевины. Сильная вакуолизация в первичной коре и сердцевине свидетельствует о наличии меристематической зоны, называемой остаточной меристемой. Она выглядит так, как если бы была остатком апикальной меристемы, сохранившимся среди более дифференцированных тканей. Ниже места закладки листового зачатка в остаточной меристеме путем продольных делений, без последующего разрастания клеток в ширину, образуются несколько вытянутые клетки первого прокамбия.
На следующих стадиях развития в остаточной меристеме дифференцируется все больше прокамбиальных тяжей. После того, как сформировались все проводящие пучки данного уровня стебля, остаточная меристема дифференцируется в межпучковую паренхиму. В узлах часть остаточной меристемы становится паренхимой листовой щели.
Прокамбий может закладываться сплошным кольцом или отдельными тяжами. Если прокамбий залегает сплошным кольцом, формируется беспучковый (непучковый) тип строения стебля. Если прокамбий закладывается отдельными тяжами, образуется пучковый тип строения стебля. Поэтому в стебле по разному размещаются первичные проводящие ткани: в первом случае сплошным кольцом, во втором – отдельными проводящими пучками, разделенными участками основной паренхимы – сердцевинными лучами.
В коллатеральных проводящих пучках первичная флоэма появляется во внешней части прокамбиального тяжа, первичная ксилема – во внутренней. Последующая дифференциация флоэмы происходит центростремительно: новые элементы флоэмы появляются ближе к центру стебля. Ксилема дифференцируется в противоположном направлении – центробежно.
Вся стела, или центральная часть стебля состоит из проводящих тканей, сердцевины (иногда она разрушается), перицикла. Наружу от перицикла размещается первичная кора. В состав первичной коры (а образуется она их периферической меристемы) входит хлоренхима, иногда секреторные элементы. Самый наружный слой периферической меристемы не образует экзодерму, как в корне, а дифференцируется в колленхиму, которая залегает или в виде тяжей или сплошным кольцом (в зависимости от типа строения стебля: пучковый или беспучковый). Самый внутренний слой первичной коры – эндодерма. Но в стебле она не выполняет регулирующей функции, как в корне. В эндодерме откладываются крахмальные зерна, и поэтому ее называют крахмалоносным влагалищем. Иногда в ней есть кристаллы оксалата кальция.
Перицикл быстро теряет меристематическую активность и преобразуется в склеренхиму, залегающую сплошным слоем или тяжами в зависимости от типа строения стебля.
Сердцевина представлена тонкостенными паренхимными клетками, в которых откладываются запасные питательные вещества. Здесь встречаются клетки-идиобласты, заполненные кристаллами, слизями и т.д. Периферическая часть сердцевины называется перимедуллярной зоной (греч. peri – вокруг, medulla – сердцевина). Клетки перимедуллярной зоны более мелкие, долго сохраняют свою жизнедеятельность.
У однодольных исходным является пучковый тип строения стебля, т.е. прокамбий залегает тяжами. Но вторичная меристема – камбий – никогда не возникает, и поэтому отсутствуют вторичные проводящие ткани. Даже при утолщении стебель образован только первичными тканями. Толстый стебель однодольных (пальмы, алоэ) развивается благодаря делению и растяжению клеток основной паренхимы. Такой рост называется диффузным вторичным ростом.
У однодольных первичная кора и центральный цилиндр не разделены четкой границей, как у двудольных, и стела подступает под эпидермис.
Для однодольных характерен диффузный тип строения стебля, когда закрытые проводящие пучки рассеяны по всему стеблю. В процессе удлинения междоузлий первичные элементы флоэмы и ксилемы (протофлоэма и протоксилема) растягиваются и разрушаются, в результате чего с ксилемной стороны пучка образуется крупная воздухоносная полость. Поэтому на лабораторных занятиях, когда мы смотрели типы пучков, на постоянном препарате поперечного среза стебля кукурузы было видно, что зрелый проводящий пучок включал два широких сосуда метаксилемы, воздушную полость, метафлоэму. Каждый пучок заключен в обкладку из склеренхимных клеток.
У однодольных часто отмирает сердцевина стебля и на ее месте образуется полость. Такой полый стебель с хорошо выраженными узлами называется соломиной. Соломина характеризуется сильным развитием механической ткани, которая образует кольцо на периферии стебля. У зрелых растений хлоренхима разрушается, и соломина приобретает золотисто-желтую окраску, свойственную склеренхиме. Вообще для однодольных характерна сильная склерификация стебля, когда до 30% всех тканей приходится на склеренхиму и может быть частичная лигнификация оболочек живых паренхимных клеток.
В стебле двудольных вторичный рост является результатом деятельности камбия. Камбий появляется в конце первого года роста побега и способствует утолщению стебля. Камбий возникает частично из прокамбия в проводящих пучках, а частично из межпучковой паренхимы. В зависимости от места образования участки камбия называются пучковым и межпучковым камбием. Будущие камбиальные клетки в межпучковых зонах детерминированы задолго до начала камбиальной деятельности. Камбий образуется сначала в клетках, прилегающих к пучку, затем распространяется вглубь сердцевинного луча. В результате межпучковый и пучковый камбий образуют в стебле сплошное камбиальное кольцо.
Различают три типа вторичного роста:
1) тип кирказон Aristolochia.
Исходным является пучковый тип строения стебля. Первичные проводящие пучки разделены широкими сердцевинными лучами. Пучковый камбий образует вторичные проводящие ткани, межпучковый – лучевую паренхиму. Такое пучковое строение сохраняется.
2) тип подсолнечник Helianthus.
Так же исходным является пучковый тип строения. Но вторичные проводящие ткани образуются как пучковым, так и межпучковым камбием. Вторичные проводящие пучки, образованые межпучковым камбием, отличаются от первичных меньшими размерами и отсутствием над пучком механической ткани перициклического происхождения. При продолжительном действии камбия вторичных пучков становится все больше. Они разрастаются, сливаются между собой, образуя сплошное кольцо вторичных проводящих тканей.
3) тип липа Tilia.
Исходным является беспучковый тип строения стебля. Общее камбиальное кольцо образует вторичные проводящие ткани. Но время от времени камбий формирует паренхимные клетки, которые в дальнейшем дифференцируются во вторичные лучи.
Строение стебля многолетних растений
У древесных и кустарниковых двудольных, а также у хвойных вторичные утолщения продолжаются много лет, и стволы у некоторых видов достигают несколько метров в диаметре. Утолщения связаны с деятельностью длительно функционирующего камбия и отчасти феллогена – двух вторичных боковых меристем. Топографически в стебле многолетнего древесного растения можно выделить три основных части: кору, древесину и сердцевину. Граница коры и древесины проходит по камбию.
Клетки камбия, в отличие от типичных меристематических клеток, всегда сильно вакуолизированы. Состояние цитоплазмы клеток камбия изменяется по сезонам.
В камбиальной зоне или камбиальном кольце встречаются две формы клеток камбия: вытянутые в вертикальном направлении веретеновидные клетки (инициали) и вытянутые в горизонтальном направлении шаровидные клетки – лучевые инициали. Если веретеновидные клетки размещаются правильными горизонтальными рядами, камбий называют ярусным. Если такого порядка в размещении веретеновидных клеток нет – камбий неярусный.
Ярусный камбий дает начало ярусной древесине и ярусной вторичной флоэме. Из неярусного камбия возникает неярусная древесина и неярусная вторичная флоэма. В эволюционном плане ярусные древесины и флоэмы считаются более высокоспециализированные, чем неярусные.
С утолщением стебля увеличивается и окружность слоев камбия. Это возможно благодаря делению камбиальных клеток в радиальном направлении.
Среди камбиальных клеток выделяют средний инициальный слой. Клетки его в результате деления формируют производные во внутреннем, а затем во внешнем направлении. За одинаковый промежуток времени во внутрь откладывается 2-3 клетки, а во внешнем направлении – только одна клетка. В результате древесины формируется больше, чем луба.
Лучевые клетки (инициали) образуют сердцевинные паренхимные лучи: первичные, которые тянутся от сердцевины к коре; и вторичные сердцевинные лучи более короткие, т.к. возникают из лучевых клеток, возникших позже.
Сердцевинные лучи могут быть однорядные и многорядные; гомогенные (т.е. состоящие из одинаковых клеток) и гетерогенные (в состав луча входят клетки, разные по строению).
Значение сердцевинных лучей: это место запасания питательных веществ (крахмала, липидов), что играет роль в период покоя (например, зимой).
Сердцевинные лучи обеспечивают радиальный перенос воды и минеральных солей из ксилемы по апопласту к камбию и вторичной флоэме, а в обратном направлении по симпласту питательных веществ.
И, наконец, по межклетникам осуществляется газообмен.
Древесина
Древесина образуется камбием и нарастает в центробежном направлении. Т.к. в нашей зоне камбий работает периодически: начинает функционировать рано весной и прекращает деятельность в конце лета, то в древесине видна концентрическая слоистость. Весной, в начале вегетационного периода, образуется ранняя древесина. Ее сосуды широкие, тонкостенные, способные проводить значительное количество воды. Вода необходима для инициации роста, особенно для увеличения в размерах новых клеток (например, клеток развивающихся листьев). Поздняя древесина содержит меньше сосудов. Они более узкие, толстостенные. Переход от ранней к поздней древесине того же года может быть постепенным. Однако граница поздней древесины одного года с ранней древесиной следующего года всегда резкая и хорошо видна на поперечном срезе. В результате образуется годовой прирост (годичное кольцо или годичный слой).
По размещению сосудов в годовом (годичном) приросте различают следующие виды древесины.
Если сосуды располагаются в основном в ранней древесине, а в поздней они отсутствуют или размещаются только мелкие сосуды, как, например, древесина дуба, ясеня, - это кольцесосудистая древесина .
Если сосуды располагаются равномерно по всему годовому приросту, хотя диаметр их в поздней древесине уменьшается, - это рассеянососудистая древесина. Встречается у березы, клена, тополя.
Есть и переходные формы между этими видами древесины.
Поскольку древесина выполняет проводящую, механическую и запасающую функции, то и в ее состав входят кроме элементов ксилемы механическая ткань в виде склеренхимы и запасающая в виде основной паренхимы. Строение всех анатомических элементов, их количество, размещение придают древесине разных растений специфические особенности, характерные для определенной таксономической группы.
Например, у более подвинутых в эволюционном плане растений есть специализированная механическая ткань (склеренхима). Примитивные в эволюционном плане двудольные (магнолиевые) не имеют древесинных волокон. Механическую роль в этом случае выполняют трахеиды.
В древесине некоторых растений есть переходные формы от трахеид к либриформу (либриформ - специализированные механические элементы древесины, состоящие из прозенхимных, заостренных на концах клеток с толстыми одревесневевшими оболочками). Встречается и перегородчатый либриформ. Его исходная веретеновидная клетка с толстыми вертикальными оболочками разделена поперек на отдельные короткие живые клетки. Такие гистологические элементы морфологически и функционально приближаются к древесной паренхиме.
Древесная паренхима выполняет запасающую и частично проводящую роль. Клетки паренхимы живые. Размещается древесная паренхима горизонтальными и вертикальными рядами. Горизонтальные ряды образуют сердцевинные лучи. Их особенности мы уже рассмотрели. Вертикальные ряды образуют саму древесную паренхиму.
Если древесная паренхима рассеяна по всему годовому приросту, это диффузная паренхима. Характерна для древесины липы, дуба, груши.
Если древесная паренхима размещается вокруг сосудов, это паратрахеальная паренхима. В древесине ясеня, клена.
По внешней границе годового прироста располагается терминальная паренхима. Пример: древесина ивы, лиственницы, магнолии.
Особенность ее строения – это отсутствие сосудов. Трахеальные элементы неперфорированы и представлены трахеидами, выполняющими и проводящую и механическую роль.
Для трахеид хвойных характерны крупные окаймленные поры с торусом. Трахеиды длинные с заостренными концами. Ранние трахеиды крупные, тонкостенные. Поздние трахеиды сплющены в радиальном направлении, толстостенные.
Мы видим, что древесина голосеменных и двудольных растений имеет существенные структурные отличия. Поэтому ее разделяют на две основные группы. Древесину голосеменных относят к мягким, а древесину двудольных – к твердым древесинам. Эти термины "твердая" и "мягкая" не обязательно сводятся только к степени плотности и твердости. Различия связаны, как мы уже сказали, со структурными особенностями.
В наклонных или изогнутых стволах возникает реактивная древесина. Ее образование вызвано тенденцией ветвей и стволов противодействовать нагрузкам, возникающим при наклонном положении, т.е. ее образование связано с процессом выпрямления этих частей растения.
Наиболее важными факторами, обусловливающими развитие реактивной древесины, являются гравитационные силы и распределение эндогенных стимуляторов роста.
Реактивная древесина отличается от обычной и анатомически и химически. Клетки реактивной древесины подвергаются в большей мере лигнификации и утолщению оболочки. У хвойных она более плотная и темная, чем окружающая ткань. Ее трахеиды короче, чем трахеиды нормальной древесины.
Радиоактивность стволовой древесины уменьшается от наружных слоев к сердцевине. Происходит некоторое увеличение загрязненности древесины от основания ствола к вершине.
Достаточно высокое содержание радиоактивных веществ в коре и побегах свидетельствует об их внешнем аэральном загрязнении.
Кора
Этот термин объединяет все ткани снаружи от камбия. Это уже вторичная кора, т.к. образуется вторичной меристемой – камбием (в отличие от первичной, которая формируется при дифференциации клеток первичной меристемы).
В состав вторичной коры входят вторичная флоэма, склеренхима (лубяные волокна и каменистые клетки), основная паренхима. Совокупность механических элементов называют твердым лубом, проводящие элементы и основная паренхима – мягкий луб.
Во вторичной коре встречаются секреторные клетки, смоляные каналы. Основная функция вторичной коры – проведение пластических веществ, также функция защиты.
На вторичную флоэму приходится значительно меньшая доля объема ствола, чем на вторичную ксилему. Это объясняется тем, что, во-первых, камбий образует меньше флоэмы, чем ксилемы, и, во-вторых, старая, нефункционирующая флоэма постепенно сминается.
В связи с увеличением окружности осевого органа паренхимные лучи расширяются и напоминают треугольник, вершина которого направлена в сторону камбия. Такая паренхима называется дилатационной (лат.dilatatio – расширение). Такое размещение паренхимы предохраняет луб от разрыва при росте стебля в толщину.
У хвойных вторичная флоэма имеет более простое строение. В лубе голосеменных гистологические элементы размещаются равномерно и, конечно, присутствуют смоляные каналы.
Все ткани снаружи от феллогена (имеется в виду самый внутренний пробковый камбий) по мере суберинизации пробковых клеток перестают получать воду и минеральные вещества. Эти все перидермы с остатками коры (отмершими клетками паренхимы и флоэмы) составляют наружную (внешнюю) кору. Живая часть коры между камбием и самым внутренним слоем феллогена называется внутренней корой.
Так, опираясь на наши знания из темы "Ткани" мы рассмотрели анатомическое строение стебля как осевого органа побега.
Сейчас рассмотрим строение бокового органа побега – листа.
ЛИСТ
Лист – главный фотосинтезирующий орган высших растений. Строение листа и его функции тесно взаимосвязаны.
Из суммарного уравнения реакций фотосинтеза:
СО 2 + Н 2 О хл-л → hv (СН 2 О) n + О 2
можно сделать вывод, что 1)листьям нужен источник СО 2 и вода; 2)листья должны быть приспособлены к поглощению солнечной энергии, у них должен быть хлорофилл; 3)как один из продуктов реакции будет выделяться О 2 ; 4)углевод должен или откладываться в запас или транспортироваться в другие части растения. Лист – весьма специализированный орган, отвечающий всем этим требованиям.
Вся эволюция листа, как вегетативного органа, проходила по пути развития приспособленности к наилучшему использованию света.
По микрофильной линии эволюции лист рассматривается как вырост телома архаичных растений типа риниофитов и сохраняет структуру строения осевого органа.
По макрофильной линии эволюции лист возник в результате сплющивания, размещения теломов в одной плоскости и последующего объединения их в одно целое. При этом была утеряна способность к длительному верхушечному нарастанию и ветвлению. Каковы же морфологические и анатомические особенности листа, возникшие в результате длительной эволюции?
Взрослый лист обычно состоит из листовой пластинки и черешка. Черешок – узкая стеблевидная часть листа между листовой пластинкой и узлом побега, при помощи которой лист ориентируется в пространстве и размещается наиболее благоприятно относительно света. Через черешок осуществляется связь листовой пластинки и стебля. В черешке хорошо развиты проводящие и механические ткани. При листопаде в черешке образуется отделительный слой.
Самая нижняя часть листа, сочлененная со стеблем, называется основой листа. Основа листа может принимать разнообразную форму. Чаще всего имеет вид небольшого утолщения и называется листовой подушечкой. Чаще при основе листа имеются разной формы и размера парные боковые выросты – прилистники. Прилистники могут быть листовидные и служить дополнительной ассимилирующей поверхностью. Прилистники могут быть игловидные и выполнять функции защиты растения.
Встречаются чешуевидные, пленчатые прилистники. В эволюционном плане наблюдается редукция прилистников и у эволюционно молодых растений они рано опадают или вообще отсутствуют.
У некоторых злаков, сельдерейных (зонтичных) основа листа разрастается и образует замкнутую или незамкнутую трубку – листовое влагалище. Листовое влагалище способствует длительному сохранению интеркалярной меристемы и служит средством дополнительной опоры побега.
Основная часть листа – листовая пластинка. В результате неравномерного освещения верхней и нижней сторон листа пластинка имеет дорзовентральное строение, т.е. строение верхней стороны листа, брюшной, отличается от строения нижней стороны, спинной. Верхняя сторона листа называется брюшной, т.к. в почке эта сторона внутренняя и повернута к оси побега. Нижняя, спинная, сторона листа в почке повернута кнаружи от растения. Таким образом, лист с дорзовентральной симметрией – двусторонний илибилатеральный или бифациальный. Характерен для двудольных.
Лист, имеющий радиальную симметрию, - эквифациальный. Такие утолщенные, цилиндрической формы листья встречаются у толстянковых. Односторонние, или изолатеральные унифациальные, листья характерны для злаков, лука.
Рассмотрим анатомическое строение листовой пластинки.
Как корень и стебель, лист состоит из покровной, проводящей и основной тканей. Т.к. лист обычно не имеет вторичного роста (за исключением незначительного роста в черешках и крупных жилках), в качестве покровной ткани у него сохраняется эпидерма.
Устьица в основном встречаются на нижней поверхности листа. В листьях двудольных устьица рассеяны по всей поверхности листа без видимого порядка. В листьях однодольных и хвойных они распределены рядами, параллельными продольной оси листа. В эпидерме некоторых злаков есть моторные или двигательные клетки. Они крупнее обычных клеток эпидермиса. Вакуоль занимает почти всю клетку. При потере тургора эти клетки сокращаются и способствуют складыванию или скручиванию листа.
Большая часть основной ткани листовой пластинки приходится на долю мезофилла. Мезофилл дифференцированный. Палисадный мезофилл размещается к верхней стороне пластинки, губчатый – к нижней. Листья с таким строением и есть бифациальные или дорзовентральные.
Если палисадный мезофилл размещается с двух сторон листа, лист унифациальный или изолатеральный.
В мезофилле злаков отчетливой дифференциации на палисадную и губчатую хлоренхиму нет. Клетки мезофилла расположены радиально вокруг пучков.
Проводящие пучки листа называются жилками, а система ветвления жилок – жилкование. Особенности расположения проводящих пучков в листовых пластинках обусловливают разные типы жилкования, которое может быть дихотомическим, параллельным, дуговидным, пальчатым и перистым.
В зависимости от того, как жилки более высоких порядков ветвления связаны между собой, отличают открытое (дихотомическое) и закрытое жилкование. При дихотомическом жилковании жилки ветвятся дихотомически, отходят под острым углом и доходят до края листовой пластинки, нигде не сливаясь с соседними жилками. При закрытом жилковании мелкие жилки в результате анастамозов (перекрещиваний) образуют сеть, которой бывает пронизана вся пластинка или большая ее часть.
Листья с пальчатым и перистым жилкованием, а это в основном двудольные, имеют среднюю, наиболее крупную жилку. Жилка состоит из первичной ксилемы и первичной флоэмы, возникающих из прокамбия и объединенных в коллатеральные пучки. Между ксилемой и флоэмой часто формируется камбий, но он не действует. Поэтому ксилема и флоэма – первичные.
Пучки образуют в листе непрерывную систему, связанную с проводящей системой стебля. Поэтому ксилема всегда ориентирована к морфологически верхней стороне листа, а флоэма – к морфологически нижней.
Средняя жилка связана латерально с уступающими ей по величине боковыми жилками. Каждая из них связана с еще более мелкими, а те в свою очередь расчленяются на еще более мелкие и т.д. Количество порядков ветвления у двудольных варьирует от 2 до 5 и более. Самые мелкие ветви образуют ячейки, внутри которых замкнуты небольшие участки мезофилла – ареолы.
В открытой дихотомической системе жилкования замкнутые ячейки отсутствуют.
В листьях однодольных (с параллельным жилкованием) продольные жилки связаны между собой более мелкими жилками – комиссуральными пучками, которые располагаются в виде простых поперечных перемычек. Количество проводящих тканей в мелких жилках постепенно уменьшается. В окончаниях пучков ксилемные элементы часто тянутся на большее растояние, чем флоэмные. Ксилема в окончаниях жилок обычно состоит из коротких трахеальных элементов, флоэма – из коротких узких ситовидных элементов и крупных клеток-спутников.
Небольшие проводящие пучки, локализованные в мезофилле, окружены одним или несколькими слоями компактно сложенных клеток, образующих обкладку пучка. Обкладки пучка могут быть паренхимными или склеренхимными. У некоторых видов наблюдается суберинизация клеточных оболочек в обкладках пучков, что свидетельствует о том, что клетки обкладки могут функционировать как эндодерма.
В крупных жилках с одной или с двух сторон находится колленхима и склеренхима. Ткань, ассоциирующаяся с крупными жилками, возвышается над поверхностью листа и образует выступы. Участок ткани, заключенный между выступами жилок, называют "интеркостальной зоной" (лат. название выступа "costa" – ребро).
У видов, имеющих толстые листья, в мезофилле встречаются различные склереиды, выполняющие функцию распорок.
Листья злаков характеризуются сильным развитием склеренхимы.
Листья голосеменных менее разнообразны по структуре, чем листья покрытосеменных. Рассмотрим на примере хвоинки сосны как приспособленность к перенесению низких температур зимой и способность к жизни в течение нескольких лет нашли свое отражение в анатомическом строении. Хвоинка имеет толстостенную эпидерму с мощной кутикулой и глубоко погруженными устьицами. Побочные клетки некоторых хвойных (Cupressaceae) имеют кутикулярный валик, который нависает над замыкающими клетками устьиц. Устьица размещаются вертикальными рядами по всем сторонам хвоинки. Под эпидермой находятся толстостенные с частично лигнифицированными оболочками клетки гиподермы. Мезофилл складчатый. В нем размещаются смоляные ходы. В центре хвоинки находятся проводящие пучки. Обычно два, реже один. Эти пучки связаны склеренхимной стяжкой. Пучки окружены трансфузионной тканью, состоящей из трахеид и паренхимных клеток. Трансфузионная ткань (лат. transfusio –– переливание) имеет отношение к транспорту воды и питательных веществ между проводящими пучками и мезофиллом.
Трансфузионная ткань окружена толстостенной эндодермой. Эндодерма часто имеет пояски Каспари на ранних стадиях развития и субериновую пластинку на более поздних. В зрелом состоянии клетки эндодермы имеют вторичные лигнифицированные оболочки.
Анатомическое строение листовой пластинки выявляет значительную пластичность. Строение изменяется в зависимости от условий существования растения и размещения листьев на растении.
Мы уже отмечали одно из направлений развития современной ботаники – экологическую анатомию растений – и говорили, что именно живое содержимое клетки – протопласт – прежде всего реагирует на изменение условий окружающей среды. Так, клетки северных растений менее вакуолизированы, имеют крупные хлоропласты. В строме выявляется много крахмальных зерен. Тилакоидная система хлоропластов развита слабее, чем у хлоропластов растений умеренных широт. Численность митохондрий у северных растений в 2-2,5 раза выше. ЭПР и аппарат Гольджи развиты сильнее. Такие особенности субмикроскопического строения клеток мезофилла растений севера характеризуются как важные адаптивные признаки, способствующие выживанию растений в суровых климатических условиях.
У растений, приспособленных к дефициту влаги, холоду отмечаются толстые клеточные оболочки, особенно в эпидерме. Сильно утолщенные наружные оболочки крупноклеточной эпидермы занимают половину полости клетки. Над эпидермой – толстый слой кутикулы. С поверхности кутикула вся покрыта толстыми чешуями воска, от чего и зависит сизоватый оттенок листьев.
Устьица погружены в углубления, высланные эпидермальными волосками.
У растений-суккулентов отмечается наличие водозапасающей ткани.
Интересные приспособления для улавливания влаги у листьев лебеды мягкой (Африка). Лист с верхней и нижней сторон покрыт сплошным слоем многорядно размещенных пузыревидных волосков. Толщина этого слоя значительно превышает толщину мезофилла и предохраняет живые ткани от высыхания. Волоски способны улавливать влагу воздуха ночью и улучшать суровый водный режим растения.
Листья и стебель лебеды раскидистой нашей лесной зоны покрыты многочисленными пузыревидными волосками, составляющими впечатление мучнистого налета, но такого плотного многорядного слоя, как у лебеды мягкой, они не образуют.
На примере сравнивания листьев двух видов лебеды видно, что систематические признаки близких таксономических единиц при расселении их в разных экологических условиях приобретают неодинаковое развитие и носят приспособительный характер.
Листья растений дождевых лесов (например, монстера) имеют продырявленные листовые пластинки и длинные черешки. Избыток воды всегда легко стекает. Устьица открыты. Палисадная ткань с крупными воздушными полостями.
Помимо температуры, влажности еще один важный абиотический фактор – свет, на который растение реагирует структурными изменениями.
Листья, развивающиеся на прямом солнечном свету, мельче теневых, но толще теневых в результате сильного развития палисадного мезофилла.
Таким образом, закрепленные в ходе эволюции приспособления к условиям среды касаются количественных отличий, но не изменяют принципов организации.
Онтогенез листа
Листья закладываются в форме небольших бугорков как боковые выросты апикальной меристемы. В образовании листового бугорка принимает участие не только туника, но и клетки корпуса. Листовой примордий разрастается из исходного выступа вверх в виде конического или игловидного выроста. Вскоре листовые зачатки окружают апикальную меристему, защищают ее и механически, и тем теплом, которое они выделяют при дыхании.
Ранний рост листа обычно разделяется на апикальный и маргинальный (краевой). Первый связан с удлинением примордия, второй – с латеральным разрастанием, в результате которого образуются две части пластинки. В соответствии с этим растущий примордий имеет апикальную меристему на верхушке и две маргинальные меристемы по бокам оси. Апикальный рост примордия не продолжительный. За ростом апекса и краев пластинки следует интеркалярный рост.
В сложном листе активность маргинальной меристемы локализуется в отдельных центрах, каждый из которых формирует отдельный листочек.
Развитие листовой пластинки происходит быстрее, чем рост черешка, который образуется позднее путем интеркалярного роста.
Если на границе между основой интеркалярной и маргинальной зон формируется перетяжка, то развивается черешковый лист. Большая часть черешка образована интеркалярной зоной. Если такой перетяжки (на границе между основой интеркалярной и маргинальной зон) не образуется, развивается сидячий лист.
В листовом зачатке однодольных растений маргинальные меристемы почти не закладываются и долгое время характерен интеркалярный рост. Отсюда основная форма листовой пластинки – линейная.
Отличия в скорости делений и растяжения клеток разных слоев пластинки приводят к образованию многочисленных межклетников и типичной структуры мезофилла.
Развитие других тканей коррелирует с развитием проводящих тканей. Прокамбий обособляется одновременно в листе и прилегающем участке конуса нарастания, образуя непрерывный тяж. Он формирует первичный проводящий пучок, который является непрерывным и общим для листа и стебля.
Онтогенез листа свидетельствует об общей природе листа и стебля как частей единого целого – побега.
С момента развертывания почки начинается внепочечная фаза развития листа. Поверхность листьев при этом увеличивается в несколько сотен, даже тысяч раз. Рост поверхности достигается за счет деления большинства клеток листа и растяжения их в длину и ширину.
Продолжительность жизни листьев зависит от генетических и климатических факторов. У листопадных деревьев и кустарников умеренного климата внепочечный период жизни листьев составляет 4-5 месяцев. Листья хвойных растений живут от двух до шести и более лет. Но в любых случаях продолжительность жизни листьев значительно меньше продолжительности жизни осевых органов растения.
Welwitschia mirabilis произрастает в каменистых пустынях юго-западной Африки. Это дерево – карлик имеет длинный корень, толстый и короткий ствол (до 50см высотой и до 1м толщиной) и два крупных листа, сохраняющиеся в течение всей жизни (до 2000 лет). Листья достигают 2-3 метра длины и постоянно нарастают у основания, отмирая у верхушки. Почти единственным источником влаги для вельвичии является густой туман, влагу которого это растение поглощает через многочисленные устьица на обеих сторонах листа (22 200 устьиц на 1см 2).
Активное отделение листьев от ветвей без повреждения ее живых тканей называется опадением. Сезонный листопад деревьев является результатом реакции на изменение длины дня. Опадение листьев осенью значительно уменьшает поверхность испарения растения, что необходимо в условиях физиологического дефицита влаги осенью и зимой. Благодаря листопаду понижается опасность полома ветвей от тяжести снега.
У большинства листьев формирование отделительного слоя происходит на протяжении онтогенеза. В этой зоне сокращается количество механической ткани. Перед опадением в трахеальных элементах образуются тилы, в ситовидных элементах откладывается каллоза. При опадении происходит ферментативное разрушение клеточных оболочек, приводящее к отделению клеток друг от друга. Изменения оболочек включают утрату цементирующей способности срединной пластинки (отчасти в результате удаления из нее кальция), гидролиз самих целлюлозных оболочек и разрыв склерифицированных трахеальных элементов.
Защитный слой, или рубец, образуется в результате отложения в оболочках и межклетниках защитных веществ, таких, как суберин или раневая камедь. У древесных растений защитный слой заменяется перидермой, которая закладывается под защитным слоем и становится непрерывной с перидермой остальной части стебля.
Сбрасывание листьев не обязательно связано с процессом растворения клеточных оболочек. Механический отрыв без предварительных химических изменений можно наблюдать при сбрасывании хвои у ели.
После опадения листьев остаются листовые рубцы со своими пучковыми рубцами. Пучковые рубцы – это обломанные концы проводящих пучков, которые шли из листовых следов в черешок листа до его опадения.
Листовые следы – это ответвления от проводящей системы стебля к листьям. Листовой след тянется от места его слияния со стеблевым пучком до выхода в лист. Один лист может иметь один или более листовых следов.
Т.к. стебель и лист имеют филогенетически общее происхождение, никакой фундаментальной разницы между листовыми следами и пучками стебля не существует. Соответствующие термины имеют описательное, топографическое значение.
В области узла, где листовой след отходит от центрального проводящего цилиндра стебля в направлении к основе листа, в цилиндре формируется паренхимная зона – листовая лакуна или листовая щель. Количество листовых следов и листовых лакун разное у разных растений.
Первичным типом узла у покрытосеменных является узел с тремя или большим количеством лакун. Основное направление эволюции покрытосеменных характеризуется уменьшением количеством лакун.
Лакуны, сердцевина, межпучковые зоны, проводящая система и расположенные на периферии проводящей системы перицикл составляют внутренний стержень осевой части растения (стебля и корня) или стелу.
Классификация стел основывается на взаимном расположении проводящих и непроводящих тканей в осевых органах на первичной стадии развития. В простейшем типе стелы, которая рассматривается как наиболее филогенетически примитивная, проводящая ткань образует сплошную колонку, причем флоэма окружает ксилему. Это протостела. Такая организация стебля биологически несовершенная, т.к. поверхность соприкосновения между проводящими и основными тканями малая.
Дальнейшая эволюция стелы шла по пути увеличения поверхности соприкосновения проводящих тканей с основными. Это достигалось двумя путями. В одном случае стела образует глубокие выросты в сторону коры и на поперечном срезе имеет форму звезды. Это актиностела.
Второй, биологически более перспективный путь развития, - это появление паренхимной сердцевины в центре стелы. Это сифоностела.
Более тесная связь между проводящими и основными тканями возникает с развитием листьев. Чем больше листьев, тем больше стела насыщена основной паренхимой. Стела приобретает вид сети (греч. сеть – dictуon). Это диктиостела.
Еще больше паренхиматизирована стела современных цветковых. Стела, представляющая собой систему проводящих тяжей и межпучковых зон, - эустела (греч. eu – хороший).
У однодольных листовые следы доходят до центра стебля, затем отклоняются к периферии. Поэтому на поперечном сечении стебля пучки кажутся разбросанными без всякого порядка. Ataktos с греч. беспорядочный. Отсюда название стелы однодольных – атактостела. Для нее характерна высокая степень паренхиматизации, когда каждый листовой след "



































- Размер: 2 Mегабайта
- Количество слайдов: 36
Описание презентации КОРЕНЬ И КОРНЕВЫЕ СИСТЕМЫ 1. Функции и эволюционное по слайдам
КОРЕНЬ И КОРНЕВЫЕ СИСТЕМЫ 1. Функции и эволюционное возникновение корня. 2. Первичное строение корня. 3. Вторичные изменения корня. 4. Образование боковых и придаточных корней. Корневые системы. 5. Специализация и видоизме-нения корней.
Корень – осевой орган, обладающий радиальной симметрией и неопределенно долго нарастающий в длину благодаря деятельности апикальной меристемы. На корне никогда не возникают листья, а апикальная меристема всегда прикрыта чехликом. Главная функция корня – поглощение воды и минеральных веществ, т. е. обеспечение почвенного питания растения. Помимо названной главной функции корни выполняют и другие функции: укрепляют растение в почве, делают возможным вертикальный рост и вынесение побегов вверх; в корнях происходит вторичный синтез различных веществ (аминокислот, алкалоидов, фитогормонов и др.); в корнях могут откладываться запасные вещества; корни взаимодействуют с корнями других растений, почвенными микроорганизмами и грибами.
Корни возникли из теломов риниофитов, распростертых по поверхности почвы. В ходе эволюции некоторые ответвления этих теломов стали углубляться в почву и дали начало корням.
Корни приспособлены к более совершенному почвенному питанию. Возникновение корней сопровождалось глубокой перестройкой всей их структуры. В них возникли специализированные ткани. Функцию поглощения веществ из почвы стали выполнять молодые окончания корней. Они сохраняют на поверхности живые клетки. Эти клетки образовали важнейшую в функциональном отношении ткань корня — ризодерму.
Функцию поглощения веществ из почвы стали выполнять молодые окончания корней. Они сохраняют на поверхности живые клетки. Эти клетки образовали важнейшую в функциональном отношении ткань корня — ризодерму. Далее в процессе эволюции происходило увеличение поглощающей поверхности корня благодаря трем факторам: 1) обильному ветвлению и образованию большого числа всасывающих окончаний; 2) постоянному нарастанию корней и передвижению всасывающих окончаний в новые участки почвы; 3) образованию корневых волосков.
Поскольку нарастание корня происходит в плотной почве, верхушечная меристема его должна быть защищена. Защиту апикальной меристемы от повреждений обеспечило появление корневого чехлика. Появление корней было вызвано нарастанием сухости климата. Наступление более засушливого климата вызвало у наземных растений необходимость прикрепления к субстрату и поглощения из него воды и питательных веществ. Однако в ходе эволюции структура корня у разных видов растений изменилась меньше, чем у стебля. Это обусловлено тем, что в почвенной среде условия более стабильны, чем в воздушной. Поэтому корень считается более «консервативным» органом, хотя он появился гораздо позже побега. Образование корней – важный ароморфоз растений. Благодаря ему растения смогли освоить более сухие почвы и формировать крупные, поднимающиеся вверх побеги.
Реакция амилопластов корневого чехлика на гравитацию. Перемещение статолитов играет важную роль в создании градиентов фитогормонов, обеспечивающих вертикальный рост корня.
Строение корня проростка пшеницы (Triticum aestivum): А — схема строения корня; Б — дифференциация клеток ризодермы и экзодермы. 1 — зона проведения, 2 — зона всасывания, 3 — зона растяжения, 4 — зона деления, 5 – корневой волосок, 6 — корневой чехлик.
Поперечный срез корня (а — однодольного, б — двудольного растения)
Первичная кора возникает из периблемы. Основную её массу составляют живые паренхимные клетки с тонкими оболочками. Между ними образуется система межклетников, вытянутых вдоль оси корня. По межклетникам циркулируют газы (СО 2). Газы необходимы для поддержания интенсивного обмена веществ в клетках коры и ризодермы. Энергичный обмен веществ в клетках коры необходим для выполнения ряда важных функций: 1) клетки коры снабжают ризодерму пластическими веществами и сами участвуют в поглощении и проведении веществ; 2) в коре синтезируются различные вещества, передающиеся затем в другие ткани; 3) в клетках коры накапливаются запасные вещества; 4) в коре часто находятся гифы грибов, образующих микоризу.
Клетки эндодермы проходят три стадии развития. В зоне всасывания эндодерма находится в первой стадии. Посредине радиальных стенок её клеток образуются пояски Каспари. Пояски Каспари перекрывают передвижение веществ через клеточные оболочки, т. е по апопласту. Вторую стадию можно наблюдать в зоне боковых корней. При этом с внутренней стороны клеточной оболочки появляется тонкая субериновая пластинка. Однако эндодерма ещё свободна пропускать растворы, так как в ней остаются отдельные пропускные клетки с тонкими стенками. Третью стадию разв и тия эндодермы можно наблюдать в зоне проведения корней однодольных. Внутренние и радиальные стенки её клеток сильно утолщаются. На поперечных срезах такие клетки имеют подковообразную форму. Пропускных клеток нет. Толстостенная эндодерма защищает проводящие ткани и повышает прочность корня.
Поясок Каспари – водонепроницаемый барьер, заставляющий воду покинуть апопласт и устремиться через мембраны клеток эндодермы в симпласт
Поступление воды из почвы в корень: вода может передвигаться по апопласту и симпласту, пока не достигнет эндодермы. Дальнейшее передвижение по апопласту невозможно.
Различные типы строения центрального цилиндра корня (первичное строение): А-диархный, Б-триархный, В- тетрархный, Г- полиархный. Типы А-В характерны для двудольных, Г – у многих однодольных. 1 — участок первичной коры, 2 — первичная флоэма, 3 — первичная ксилема.
Можно выделить 4 этапа перехода корня от первичного строения ко вторичному: 1) появление камбия между участками первичной флоэмы и ксилемы; 2) образования феллогена перициклом; 3) сбрасывание первичной коры; 4) смена радиального расположения проводящих тканей коллатеральным.
Переход от первичного строения корня к вторичному 1 -первичная флоэма, 2 -первичная ксилема, 3 — камбий, 4 — перицикл, 5 — эндодерма, 6 — мезодерма, 7 — ризодерма, 8 — экзодерма, 9 -вторичная ксилема, 10 — вторичная флоэма, 11 — вторичная кора, 12 — феллоген, 13 — феллема.
Схема первичной дифференциации проводящих тканей в корне гороха 1 -центростремительная дифференциация ксилемы, 2 -эпидерма, 3 -первичная кора, 4 -эндодерма, 5 -первые дифференцированные элементы ксилемы, 6 -недифференцированные элементы ксилемы, 7 -первые дифференцированные элементы флоэмы, 9 -апикальная меристема, 10 -корневой чехлик.
Вторичные изменения корня однодольных. Подавляющее большинство однодольных растений первичное строение корня сохраняет до конца жизни. Однако при этом многие элементы корня подвергаются одревеснению. У древесных однодольных (пальмы, драцены, юкки) в коре корня из клеток паренхимы или из перицикла возникает слой меристемы. Из него формируются ряды закрытых проводящих пучков. Вслед за этим рядом проводящих пучков в периферической части паренхимы первичной коры появляется новый слой образовательной ткани. Этот слой меристемы дает начало новому ряду проводящих пучков. Таким образом происходит утолщение корня.
Придаточные корни возникают на различных органах растений – на стеблях, листьях и корнях. Придаточные корни, возникшие на стебле, называются стеблеродные, возникшие на корне – корнеродные. Боковые и придаточные корни имеют эндогенное происхождение, т. е. закладываются во внутренних тканях.
Заложение бокового корня начинается с деления клеток перицикла. При этом на поверхности стелы образуется меристематический бугорок. После ряда делений клеток меристематического бугорка возникает боковой корешок. Он имеет собственную апикальную меристему и чехлик. Зачаток бокового корня растет, прорывает первичную кору материнского корня и выдвигается наружу. Обычно боковые корни возникают против элементов ксилемы. Поэтому они располагаются правильными продольными рядами вдоль корня. Возникают они в зоне поглощения или несколько выше. Заложение боковых корней происходит акропетально, т. е. от основания корня к его верхушке.
Придаточные корни обычно закладываются в тканях, способных к меристематической активности: в перицикле, камбии, феллогене. Эндогенное образование боковых (и придаточных) корней имеет приспособительное значение. Если бы ветвление происходило в апексе, то продвижение корня в почве было затруднено.
Дихотомическое ветвление в корневой системе плауна булавовидного (Lycopodium clavatum) 1 – изотомное дихотомическое ветвление самых тонких корешков
Микоризы: А – эктотрофная микориза дуба, Б, В – эндотрофная микоризу ятрышника. Клубеньки на корнях люпина
Функции корня. Корень - основной орган высшего растения. Функции корней следующие:
Всасывают из почвы воду и растворенные в ней минеральные соли, транспортирует их вверх по стеблю, листьям и репродуктивным органам. Функцию всасывания выполняют корневые волоски (или микоризы), расположенные в зоне всасывания.
Благодаря большой прочности закрепляют растение в почве.
- При взаимодействии воды, ионов минеральных солей и продуктов фотосинтеза синтезируют продукты первичного и вторичного метаболизма.
- Под действием корневого давления и транспирации ионы водных растворов минеральных веществ и органические вещества по сосудам ксилемы корня передвигаются по восходящему току в стебель и листья.
- В корнях откладываются в запас питательные вещества (крахмал, инулин и др.).
- В корнях осуществляется биосинтез вторичных метаболитов (алкалоиды, гормоны и другие БАВ).
- Синтезированные в меристематических зонах корней ростовые вещества (гиббереллины и др.) необходимы для роста и развития надземных частей растения.
- За счет корней осуществляется симбиоз с почвенными микроорганизмами - бактериями и грибами.
- С помощью корней происходит вегетативное размножение многих растений.
10. Некоторые корни выполняют функцию дыхательного органа (монстера, филодендрон и др.).
11. Корни ряда растений выполняют функцию «ходульных» корней (фикус баньян, панданус и др.).
12. Корень способен к метаморфозам (утолщения главного корня образуют «корнеплоды» у моркови, петрушки и др.; утолщения боковых или придаточных корней образуют корневые клубни у георгин, земляных орешков, чистяка и др., укорачивание корней у луковичных растений).
Корень - осевой орган, обычно цилиндрической формы, с радиальной симметрией, обладающий геотропизмом. Растет до тех пор, пока сохраняется верхушечная меристема, покрытая корневым чехликом. На корне в отличие от побега никогда не образуются листья, зато, как и побег, корень ветвится, образуя корневую систему.
Корневая система - это совокупность корней одного растения. Характер корневой системы зависит от соотношения роста главного, боковых и придаточных корней.
^ Типы корней и корневых систем. В зародыше семени все органы растения находятся в зачаточном состоянии. Главный, или первый, корень развивается из зародышевого корешка. Главный корень расположен в центре всей корневой системы, стебель служит продолжением корня, и вместе они составляют ось первого порядка. Участок на границе между главным корнем и стеблем называется корневой шейкой. Этот переход от стебля к корню заметен по разной толщине стебля и корня: стебель более толстый, чем корень. Участок стебля от корневой шейки до первых зародышевых листьев - семядолей называютподсемядольным коленом или гипокотилем . От главного корня в стороны отходят боковые корни очередных порядков. Такая корневая система называетсястержневой ,у многих двудольных растений она способна к ветвлению. Ветвистая корневая система - разновидность стержневой корневой системы. Боковое ветвление корня характеризуется тем, что новые корни закладываются на некотором расстоянии от верхушки и образуются эндогенно - во внутренних тканях материнского корня предшествующего порядка за счет активности перицикла. Чем больше отходит боковых корней от главного корня, тем больше площадь питания растения, поэтому существуют специальные агротехнические приемы, усиливающие способность главного корня образовывать боковые, например прищипывание или пикирование главного корня на l/3 его длины. После пикирования некоторое время главный корень перестает расти в длину, в то время как боковые корни интенсивно растут.
У двудольных растений главный корень, как правило, сохраняется всю жизнь, у однодольных зародышевый корешок быстро отмирает, главный корень не развивается, а от основания побега образуются придаточные корни,которые также имеют ответвления первого, второго и т.д. порядков. Такая корневая система называется мочковатой. Придаточные корни, как и боковые, закладываются эндогенно. Они могут образовываться на стеблях и листьях. Способность растений развивать придаточные корни широко используется в растениеводстве при вегетативном размножении растений (размножение стеблевыми и листовыми черенками). Надземными стеблевыми черенками размножают иву, тополь, клен, смородину черную и др.; листовыми черенками - фиалку узамбарскую, или сенполию, некоторые виды бегоний. Подземными черенками видоизмененных побегов (корневищ) размножают многие лекарственные растения, например, ландыш майский, купену лекарственную и др. Некоторые растения образуют много придаточных корней при окучивании нижней части стебля (картофель, капуста, кукуруза и др.), создавая тем самым дополнительное питание.
У высших споровых растений (плауны, хвощи, папоротники) главный корень вообще не возникает, у них формируются только придаточные корни, отходящие от корневища. У многих двудольных травянистых корневищных растений главный корень часто отмирает и преобладает система придаточных корней, отходящих от корневищ (сныть, крапива, лютик ползучий и др.).
По глубине проникновения в почву первое место принадлежит стержневой корневой системе: рекордная глубина проникновения корней, по некоторым сведениям, достигает 120 м! Однако мочковатая корневая система, обладая в основном поверхностным расположением корней, способствует созданию дернового покрова и предупреждает эрозию почвы.
Общая длина корней в корневой системе различна, некоторые корни достигают нескольких десятков и даже сотен километров. Например, у пшеницы длина всех корневых волосков достигает 20 км, а у озимой ржи общая длина корней первого, второго и третьего порядков свыше 180 км, а с присоединением корней четвертого порядка - 623 км. Несмотря на то, что корень растет всю жизнь, рост его ограничен влиянием корней других растений.
Степень развития корневых систем на разных почвах в разных природных зонах неодинакова. Так, в песчаных пустынях, где глубоко залегают грунтовые воды, корни некоторых растений уходят на глубину 40 м и более (злак селин, просопис сережкоцветный из семейства Бобовые и др.). Растения-эфемеры полупустынь имеют поверхностную корневую систему, которая приспособлена к быстрому впитыванию ранневесенней влаги, вполне достаточной для быстрого прохождения всех фаз вегетации растений. На глинистых, плохо аэрируемых подзолах таежной лесной зоны корневая система растений на 90 % сосредоточена в поверхностном слое почвы (10-15 см), растения имеют «питающие корни» (ель европейская). Например, у саксаула корни в разное время года используют влагу разных горизонтов.
Очень важный фактор в распределении корневой системы - влажность. Направление корней идет в сторону большей влажности, однако в воде и в переувлажненной почве корни ветвятся намного слабее.
Степень развития корневых систем, глубина проникновения корней и другие пластические характеристики корня зависят от внешних условий и в то же время наследственно закреплены за каждым видом растений.
^ Зоны молодого корня. В молодом корне различают: 1) зону деления, прикрытую корневым чехликом; 2) зону растяжения клеток, или зону роста; 3) зону всасывания, или зону корневых волосков; 4) проводящую зону.
^ Зона деления представляет собой кончик корня, снаружи покрытый корневым чехликом, защищающим верхушечную, или апикальную, меристему. На ощупь молодой кончик корня скользкий из-за выделяемой клетками слизи. При росте корня в длину слизь снижает трение кончика корня о почву. По выражению академика В.Л. Комарова, корневой чехлик «роет землю», он защищает делящиеся клетки меристемы от механических повреждений, а также контролируетположительный геотропизм самого корня, т. е. способствует росту корня и проникновению его в глубь почвы. Корневой чехлик состоит из живых паренхимных клеток, в которых присутствуют крахмальные зерна. Под чехликом находится зона деления, или конус нарастания корня, представленный первичной образовательной тканью (меристемой). В результате активного деления верхушечной меристемы корня формируются все прочие зоны и ткани корня. Зона деления молодого корня составляет в длину всего лишь 1 мм. От других зон внешне она отличается желтым цветом.
^ Зона растяжения, или зона роста, протяженностью в несколько миллиметров внешне прозрачна, состоит из практически неделящихся, но растягивающихся в продольном направлении клеток. Клетки увеличиваются в размерах, в них появляются вакуоли. Клетки характеризуются высоким тургором. В зоне растяжения происходит дифференциация первичных проводящих тканей и начинают образовываться постоянные ткани корня.
Выше зоны растяжения расположена зона всасывания. Ее протяженность 5 - 20 мм. Зона всасывания представлена корневыми волосками - выростами клеток эпидермы. С помощью корневых волосков происходит всасывание из почвы воды и растворов солей. Чем многочисленнее корневые волоски, тем больше всасывающая поверхность корня. На 1 мм у поверхности корня может быть расположено около 400 корневых волосков. Корневые волоски недолговечны, живут 10 - 20 дней, после чего отмирают. Длина корневых волосков у разных растений от 0,5 - 1,0 см. Молодые корневые волоски образуются над зоной растяжения, а отмирают над зоной всасывания, поэтому зона корневых волосков постоянно перемещается по мере роста корня и растение получает возможность поглощать воду и растворенные в ней питательные вещества из разных почвенных горизонтов.
Выше зоны всасывания начинается зона проведения, или зона боковых корней. Поглощенная корнем вода и растворы солей транспортируются по сосудам древесины вверх к надземным частям растения.
Между зонами корня нет резких границ, а наблюдается постепенный переход.
6. Метаморфозы корня. Их биологическое значение. Микориза. У большинства растений в одной и той же корневой системе отчетливо различаются ростовые и сосущие окончания. Ростовыеокончания обычно более мощные, быстро удлиняются и продвигаются вглубь почвы. Зона растяжения у них хорошо выражена, и апикальные меристемы работают энергично. Сосущие окончания, возникающие в большом количестве на ростовых корнях, удлиняются медленно, и их апикальные меристемы почти перестают работать. Сосущие окончания как бы останавливаются в почве и интенсивно ее «обсасывают».
У древесных растений различают толстые скелетные и полускелетные корни, на которых образуются недолговечные корневые мочки . В состав корневых мочек, непрерывно заменяющих друг друга, входят ростовые и сосущие окончания.
Если корни выполняют особые функции, их строение меняется. Резкое, наследственно закрепленное видоизменение органа, вызванное сменой функций, носит название метаморфоза . Видоизменения корней очень разнообразны.
Корни многих растений образуют симбиоз с гифами почвенных грибов, называемый микоризой («грибокорень»). Микориза образуется на сосущих корнях в зоне поглощения. Грибной компонент облегчает корням получение воды и минеральных элементов из почвы, часто гифы грибов заменяют корневые волоски. В свою очередь, гриб получает от растения углеводы и другие питательные вещества. Различают два основных типа микоризы. Гифы эктотрофной микоризы образуют чехол, окутывающий корень снаружи. Эктомикориза широко распространена у деревьев и кустарников. Эндотрофная микориза встречается в основном у травянистых растений. Эндомикориза находится внутри корня, гифы внедряются в клетки коровой паренхимы. Микотрофное питание очень широко распространено. Некоторые растения, например орхидные, вообще не могут существовать без симбиоза с грибами.
На корнях бобовых возникают особые образования – клубеньки , в которых поселяются бактерии из рода Rhizobium. Эти микроорганизмы способны усваивать атмосферный молекулярный азот, переводя его в связанное состояние. Часть веществ, синтезированных в клубеньках, усваивают растения, бактерии, в свою очередь, используют вещества, находящиеся в корнях. Этот симбиоз имеет большое значение для сельского хозяйства. Бобовые растения благодаря дополнительному источнику азота богаты белками. Они дают ценные пищевые и кормовые продукты и обогащают почву азотистыми веществами.
Очень широко распространены запасающие корни. Они обычно утолщены и сильно паренхиматизированы. Сильно утолщенные придаточные корни называют корневыми шишками , или корнеклубнями (георгин, некоторые орхидные). У многих, чаще двулетних, растений со стержневой корневой системой возникает образование, носящее название корнеплода . В образовании корнеплода принимают участие и главный корень, и нижняя часть стебля. У моркови почти весь корнеплод составлен корнем, у репы корень образует лишь самую нижнюю часть корнеплода (рис. 4.12).
Корнеплоды культурных растений возникли в результате длительного отбора. В корнеплодах сильно развита запасающая паренхима и исчезли механические ткани. У моркови, петрушки и других зонтичных паренхима сильно развита во флоэме; у репы, редьки и других крестоцветных – в ксилеме. У свеклы запасные вещества откладываются в паренхиме, образованной деятельностью нескольких добавочных слоев камбия (рис. 4.12 ).
У многих луковичных и корневищных растений образуются втягивающие , или контрактильные корни (рис. 4.13, 1 ). Они могут укорачиваться и втягивать побег в почву на оптимальную глубину на время летней засухи или зимних морозов. Втягивающие корни имеют утолщенные основания с поперечной морщинистостью.
Дыхательные корни, или пневматофоры (рис. 4.13, 2 ) образуются у некоторых тропических древесных растений, живущих в условиях недостатка кислорода (таксодиум, или болотный кипарис; растения мангровых зарослей, обитающие по болотистым берегам океанических побережий). Пневматофоры растут вертикально вверх и высовываются над поверхностью почвы. Через систему отверстий в этих корнях, связанных с аэренхимой, воздух поступает в подводные органы.
У некоторых растений для поддержания побегов в воздушной среде образуются дополнительные опорные корни. Они отходят от горизонтальных ветвей кроны и, достигнув поверхности почвы, интенсивно ветвятся, превращаясь в столбовидные образования, поддерживающие крону дерева (столбовидные корни баньяна) (рис. 4.15, 2). Ходульные корни отходят от нижних участков стебля, придавая стеблю устойчивость. Они образуются у растений мангровых зарослей, растительных сообществ, развивающихся на затопляемых во время прилива тропических берегах океанов (рис. 4.15, 3 ), а также у кукурузы (рис. 4.15, 1 ). У фикуса каучуконосного образуются досковидные корни. В отличие от столбовидных и ходульных, они являются по происхождению не придаточными, а боковыми корнями.
Рис. 4.15. ^ Опорные корни : 1 – ходульные корни кукурузы; 2 – столбовидные корни баньяна; 3 – ходульные корни ризофоры (пр – зона прилива; от – зона отлива; ил – поверхность илистого дна).
Понятие о побеге. Морфологическое расчленение побега. Узлы и междузлия. Верхушечный рост побега. Строение и деятельностьконусанарастания. Побегом называется стебель с расположенными на нем листьями и почками.
Участки стебля, на которых развиваются листья,называют узлами.
Участки стебля между двумя ближайшими узламиназывают междоузлиями.
Угол между листом и находящимся выше междоузлием называется пазухой листа.
В пазухе листа образуется пазушная почка. Побег состоит из повторяющихся участков - метамеров
.
Один метамер включает междоузлие, узел, лист и пазушную почку. Побегом называют комплекс, состоящий из стебля и листьев. Первичный побег закладывается в зародыше, где он представлен почечкой. Почечка состоит из зародышевого стебелька - эпикотиля, апикальной меристемы и одного или нескольких листовых примордиев (зачатков листьев). При прорастании семени стебелек удлиняется. Из апикальной меристемы развиваются новые листовые примордии, из листовых примордиев
развиваются листья, а впазухах листьев
формируются примордии почек. Этот алгоритм развития в ходе образования побеговой системы растения многократно может повторяться.
У сформировавшегося побега различают узлы - часть побега, где лист соединяется со стеблем; междоузлия - часть побега между узлами, обычно это часть стебля; листовые пазухи - угол между листом и восходящей частью стебля.
Частью побега являются и почки. Это, прежде всего, верхушечная почка, представляющая конус нарастания побега. В пазухах листьев у семенных растений образуются пазушные, или боковые почки. Если они развиваются одна над другой (жимолость, грецкий орех, робиния и др.), то называются сериальными. Если почки развиваются в пазухах листьев рядом, друг около друга (слива, злаки и пр.), то они называются коллатеральными. Почки могут образовываться эндогенно в районе междоузлий. Эти почки называются придаточными.
У деревьев и кустарников холодного и умеренного климата образуются зимующие, или покоящиеся почки, которые часто называют глазками. Из этих почек на следующий год развиваются новые побеги. Наружные листья у этих почек обычно превращаются в почечные чешуи, защищающие внутренние части почки от повреждения.
Зимующие, или покоящиеся почки образуются и у многолетних трав, на тех органах, которые не отмирают на зиму, т.е. на корневищах, у основания стеблей и т.п. Эти почки называют почками возобновления. Из них весной развиваются надземные побеги.
Все вышеперечисленные почки называются вегетативными. Такие почки состоят из апекса, зачаточных узлов, зачаточных междоузлий,листовых примордиев , выше которых могут развиваться почечные примордии, и зачаточных листьев.
Из почки, не имеющей почечных примордиев, развивается простой или неветвистый побег . Из почки с почечными примордиями развивается ветвистый побег .
Кроме того, у семенных растений бывают еще и генеративные почки. Это цветковые почки и почки, дающие начало шишкам голосеменных. Они отличаются от вегетативных по внешнему виду. Кроме апекса, зачаточных междоузлий и зачаточных узлов, такие почки имеют примордии, которые дают начало частям цветка или частям шишек. У почек, дающих начало соцветиям, образуются цветковые примордии.
Наконец, есть, так называемые, смешанные почки, из которых образуются облиственные побеги с цветками.
Морфологическая характеристика побега подразумевает описание строения узлов, междоузлий, почек. Обязательно указывается тип листорасположения. У большинства растений он является очередным - в узле расположен один лист, но может быть супротивным или мутовчатым. Определенный тип листорасположения формирует листовую мозаику, что позволяет наилучшим образом использовать пространство для обеспечения равномерного освещения листа.
С процессом роста и развития побега связано и деление листьев на три категории: низовые листья, срединные листья , верхушечные, или верховые листья. При морфологическом описании листьев обычно описывают срединные листья, но полное морфологическое описание требует отдельного описания всех категорий листьев, т.к. даже срединные листья на одном побеге имеют различия. Это явление носит название гетерофиллии или разнолистности.
Верхушечный рост побега - рост побега в длину за счет видоизменения конуса нарастания, заложения и роста в его основании зачаточных листьев. В процессе видоизменения конус нарастания увеличивается в длину, усложняется и изменяет свою форму.
Почка . Это зачаточный побег. Он состоит из меристематической оси, заканчивающейся конусом нарастания (зачаточный стебель), и листовых примордиев (зачаточных листьев), то есть из серии зачаточных метамеров. Расположенные ниже дифференцированные листья прикрывают конус нарастания и примордии. Так устроена вегетативная почка. У вегетативно-репродуктивной почки конус нарастания превращен в зачаточный цветок или зачаточное соцветие. Репродуктивные (цветочные) почки состоят только из зачаточного цветка или соцветия и не имеют зачатков фотосинтезирующих листьев.
13. Метаморфизированные побеги.
Возникновение их часто связано с выполнением функций вместилища запасных продуктов, перенесения неблагоприятных условий года, вегетативного размножения.
Корневище - это многолетний подземный побег с горизонтальным, восходящим или вертикальным направлением роста, выполняющий функции накопления запасных продуктов, возобновления, вегетативного размножения. Корневище имеет редуцированные листья в виде чешуек, почки, придаточные корни. Запасные продукты накапливаются в стеблевой части. Нарастание и ветвление происходит так же, как у обычного побега. Корневище отличают от корня по наличию листьев и отсутствию корневого чехлика на верхушке. Корневище может быть длинным и тонким (пырей) или коротким и толстым. Ежегодно из верхушечной и пазушных почек образуются надземные однолетние побеги. Старые части корневища постепенно отмирают. Растения с горизонтальными длинными корневищами, формирующими много надземных побегов, быстро занимают большую площадь, и если это сорняки (пырей), то борьба с ними довольно затруднительна. Такие растения используют для закрепления песков (колосняк, аристида). В луговодстве злаки с длинными горизонтальными корневищами называют корневищными (полевица, мятлик), а с короткими - кустовыми (тимофеевка, белоус). Корневища встречаются в основном у многолетних травянистых растений, но иногда у кустарников (бересклет) и кустарничков (брусника, черника).
Клубень - это утолщенная часть побега, вместилище запасных продуктов. Клубни бывают надземными и подземными.
Надземный клубень представляет собой утолщение главного (кольраби) или бокового (тропические орхидеи) побега и несет нормальные листья.
Подземный клубень - утолщение гипокотиля (цикламен) или недолговечного подземного побега - столона (картофель). Листья на подземном клубне редуцируются, в пазухах их находятся почки, называемые глазками.
Надземный столон - это недолговечный ползучий побег, служащий для распространения (захвата территории) и вегетативного размножения. Он имеет длинные междоузлия и зеленые листья. На узлах образуются придаточные корни, а из верхушечной почки - укороченный побег (розетка), который после отмирания столона продолжает самостоятельное существование. Нарастает надземный столон симподиалыю. Надземные столоны, утратившие функцию фотосинтеза и выполняющие в основном функцию вегетативного размножения, иногда называют усами (земляника).
Луковица - это укороченный стебель (донце), несущий многочисленные, тесно сближенные листья и придаточные корни. На верхушке донца находится почка. У многих растений (лук, тюльпан, гиацинт и др.) из этой почки образуется надземный побег, а из боковой пазушной почки формируется новая луковица. Наружные чешуи в большинстве случаев сухие, пленчатые и выполняют защитную функцию, внутренние - мясистые, заполненные запасными продуктами. По форме луковицы бывают шаровидные, яйцевидные, сплющенные и т. д.
Клубнелуковица внешне похожа на луковицу, но все листовые чешуи у нее сухие, а запасные продукты откладываются в стеблевой части (шафран, гладиолус).
Колючки имеют различное происхождение - из побега (яблоня, груша, терн, боярышник, гледичия, цитрусовые), листа (барбарис) или его частей: рахиса (астрагал), прилистников (акация белая), участка пластинки (сложноцветные). Колючки характерны для растений жарких сухих местообитаний.
Усики образуются из побега (виноград), листа или его частей: рахиса и нескольких листочков (горох), пластинки (чина.), прилистников (сассапариль). Служат для прикрепления к опоре.
Филлокладии - это плоские листовидные побеги, расположенные в пазухах редуцированных листьев. На них образуются цветки. Встречаются у растений преимущественно засушливых местообитаний (иглица, филлантус). Ловчие аппараты - видоизмененные листья, свойственные насекомоядным.растениям (росянка, мухоловка). Имеют форму кувшинчиков, урночек, пузырьков или же захлопывающихся и завертывающихся пластинок. Небольшие насекомые, попадая в них, погибают, растворяются при помощи ферментов и потребляются растениями в качестве главным образом дополнительного источника минеральных веществ.
Корень - подземный осевой элемент растений, являющийся самой важной их частью, их основным вегетативным органом. Благодаря корню растение закрепляется в почве и удерживается там в течение всего жизненного цикла, а также обеспечивается водой, минеральными и питательными веществами, содержащимися в ней. Существуют разные виды и типы корней. Каждый из них имеет свои отличительные характеристики. В этой статье мы рассмотрим существующие виды корней, типы корневых систем. Также познакомимся с их характерными особенностями.
Какие есть виды корней?
Стандартный корень характеризуется нитевидной либо узкоцилиндрической формой. У многих растений, помимо основного (главного) корня, развиты еще и другие виды корней - боковые и придаточные. Рассмотрим подробнее, что они собой представляют.
Главный корень
Этот растительный орган развивается из зародышевого корешка семени. Главный корень всегда один (другие виды корней растений обычно присутствуют во множественном числе). Сохраняется он у растения на протяжении всего жизненного цикла.
Для корня характерен положительный геотропизм, то есть за счет земного притяжения он углубляется в субстрат вертикально вниз.
Придаточные корни
Придаточными называют виды корней растений, которые образуются на других их органах. Этими органами могут быть стебли, листья, побеги и пр. К примеру, у злаков имеются так называемые первичные придаточные корни, которые заложены еще в стебельке зародыша семени. Развиваются они в процессе прорастания семени практически одновременно с основным корнем.
Также встречаются листовые придаточные виды корней (образующиеся в результате укоренения листьев), стеблевые или узловые (образующиеся из корневищ, надземных или подземных стеблевых узлов) и пр. На нижних узлах образуются мощные корни, которые носят название воздушных (либо опорных).
Появление придаточных корней обуславливает способность растения к вегетативному размножению.
Боковые корни
Боковыми называют корни, возникающие в качестве бокового ответвления. Могут образовываться как на главном, так и на придаточных корнях. Кроме того, могут ответвляться и от боковых, в результате чего образуются боковые корни более высоких порядков (первого, второго и третьего).
Крупные боковые органы характеризуются поперечным геотропизмом, то есть их рост происходит в практически горизонтальном положении либо под углом к почвенной поверхности.
Что называется корневой системой?
Корневой системой называют все виды и типы корней, имеющихся у одного растения (то есть их совокупность). В зависимости от соотношения роста главного, боковых и придаточных корней определяется ее вид и характер.
Типы корневых систем
Если главный корень развит очень хорошо и заметен среди корней другого вида, это означает, что растение имеет стержневую систему. Присуща она в основном двудольным растениям.
Корневая система этого типа отличается глубоким прорастанием в почву. Так, например, корни некоторых трав могут проникать на глубину 10-12 метров (осот, люцерна посевная). Глубина проникновения корней деревьев в некоторых случаях может достигать 20 м.
Если же значительнее выражены придаточные корни, развивающиеся в большом количестве, а основной характеризуется медленным ростом, то образуется корневая система, которую называют мочковатой.

Такой системой характеризуются, как правило, и некоторые из травянистых. Несмотря на то что корни мочковатой системы проникают не так глубоко, как у стержневой, они лучше оплетают частицы грунта, прилегающие к ним. Многие рыхлокустовые и корневищные злаковые травы, образующие обильное количество мочковатых тонких корней, широко используются для закрепления оврагов, грунтов на откосах и пр. В число наилучших трав-задернителей входят костер безостый, овсяница, и др.

Видоизмененные корни
Кроме типичных, охарактеризованных выше, существуют другие виды корней и корневых систем. Они называются видоизмененными.
Запасающие корни
К запасающим относятся корнеплоды и корнеклубни.
Корнеплод представляет собой утолщение основного корня в связи с отложением в нем питательных веществ. Также в образовании корнеплода участвует нижняя часть стебля. Состоит в большей степени из запасающей основной ткани. Примерами корнеплодов могут служить петрушка, редис, морковь, свекла и др.

Если утолщенными запасающими корнями являются боковые и придаточные корни, то они называются корневыми клубнями (шишками). Они развиты у картофеля, батата, георгина и пр.
Воздушные корни
Это боковые корни, растущие в надземной части. Присутствуют у ряда тропических растений. Воду и кислород поглощают из воздушной среды. Имеются у тропических растений, произрастающих в условиях недостатка минеральных веществ.

Дыхательные корни
Это разновидность боковых корней, которые растут вверх, поднимаясь над поверхностью субстрата, воды. Такие виды корней образуются у растений, произрастающих на слишком увлажненных почвах, в условиях болот. При помощи таких корней растительность получает из воздуха недостающий кислород.
Опорные (досковидные) корни
Эти виды корней деревьев характерны для крупных пород (бук, вяз, тополь, тропические и т. п.) Представляют собой треугольные вертикальные выросты, образованные боковыми корнями и проходящие у поверхности почвы либо над ней. Их называют еще досковидными, поскольку они напоминают доски, которые прислонены к дереву.
Корни-присоски (гаустории)
Это вид дополнительных придаточных корней, развивающихся на стебле вьющихся растений. С их помощью растения имеют возможность прикрепляться к определенной опоре и подниматься (плестись) вверх. Такие корни имеются, к примеру, у фикуса цепкого, плюща и т. п.

Втяжные (контрактильные) корни
Характерны для растений, корень которых резко сокращается в продольном направлении у основания. Примером могут служить растения, имеющие луковицы. Втяжные корни обеспечивают луковицам и корнеплодам некоторое углубление в почву. Кроме того, их наличием обуславливается плотное прилегание розеток (например, у одуванчика) к земле, а также подземное положение вертикального корневища и корневой шейки.
Микориза (грибокорень)
Микоризой называют симбиоз (взаимовыгодное сожительство) корней высших растений с грибными гифами, которые оплетают их, выполняя функции корневых волосков. Грибы обеспечивают растения водой и питательными веществами, растворенными в ней. Растения, в свою очередь, дают грибам необходимые для их жизнедеятельности органические вещества.
Присуща микориза корням многих высших растений, в особенности древесных.
Бактериальные клубеньки
Это видоизмененные боковые корни, которые приспособлены к симбиотическому сожительству с фиксирующими азот бактериями. Образование клубеньков происходит за счет проникновения внутрь молодых корней. Такое взаимовыгодное сожительство позволяет растениям получать азот, который бактерии переводят из воздуха в доступную для них форму. Бактериям же предоставляется особое местообитание, где они могут функционировать, не конкурируя с другими видами бактерий. Кроме того, они используют вещества, присутствующие в корнях растительности.

Характерны бактериальные клубеньки для растений семейства Бобовых, которые широко используются в качестве мелиорантов в севооборотах с целью обогащения почв азотом. Наилучшими азотфиксирующими растениями считаются стержнекорневые бобовые травы, такие как люцерна синяя и желтая, красный и эспарцет, лядвенец рогатый и пр.
Кроме вышеперечисленных метаморфоз, существуют другие виды корней, такие как корни-подпорки (способствуют укреплению стебля), ходульные корни (помогают растениям не тонуть в жидкой грязи) и корневые отпрыски (имеют придаточные почки и обеспечивают вегетативное размножение).









